
Section 3 The birth of life
Introduction
About 4 billion years ago, a large-scale primordial ocean was formed, and the stage was finally set for the “birth of life.”
Not inferior to the “birth of the universe,” which is the transition from “nothing to being” and the discontinuous manifestation, “the birth of life” is a dramatic event that can be compared to it.
In the previous section, we looked at the events of “the birth of the solar system” and “the birth of the earth”. In fact, these are innovative events in the sense that they create something that has never existed before. However, the story of its birth is not a discontinuous manifestation of the level of “nothing to being,” but is realized from the integral accumulation of continuous phenomena under a specific environment.
It is becoming clear that the birth of life is not a discontinuous manifestation, but an integral accumulation of continuous phenomena under a specific environment. Not only that, but recent excellent research is revealing:
- It means that life is deeply related to the “flow of energy”. In other words, “energy is the cornerstone of the evolution of life,” and the characteristics of life evolution can be understood only when energy is brought into the equation. Moreover, it is not only evolution, but this relationship between energy and life goes back to the beginning of life.
- The fundamental nature of “life” is an inevitable manifestation of the imbalances that have arisen on the planets that are constantly operating. Life was born through the flow of energy, and the “proton gradient” was the key to the emergence of cells.
Let’s take a look at how this is done below.
Chapter 1 Life
What is life?
The novel coronavirus disease, which became a pandemic in 2020, spread widely. Influenza appears in the winter following the new coronavirus infection. “Viruses” such as norovirus, rubella virus, and hepatitis virus are troublesome entities that cause various infectious diseases. Is this nasty virus a living organism?
In fact, viruses are not living organisms. This is because its own “metabolism” is not working. It is totally dependent on the power of the host (Note 15).
[15] Currently, many biologists accept the definition of an organism as one that satisfies the following three conditions: (1) it is separated from the outside world by a membrane, (2) it is metabolized (the flow of matter and energy), and (3) it makes a copy of itself. This is a surprisingly simple definition, and it may seem strange that this is the only way to define an organism. However, at present, only living organisms meet all three conditions.
Here, the important concept of “metabolism” has appeared. “Metabolism” refers to a series of syntheses and chemical reactions carried out by an organism to sustain life, using inorganic substances and organic compounds taken in from the external environment as raw materials. It is an abbreviation of “renewal metabolism”. Through these pathways, organisms enable their growth and reproduction, maintaining their systems.
“Metabolism” can be broadly divided into two categories: “catabolism” and “assimilation”.
- “Catabolism” is the process of obtaining energy by decomposing a substance, for example, “cellular respiration” corresponds to this.
- “Assimilation” is the process of synthesizing substances using energy, such as “synthesis of proteins, nucleic acids, polysaccharides, and lipids”.
However, is the activity of “metabolism” really an indispensable property for life? Is it the element that defines life and clearly distinguishes animate and inanimate objects?
Do we need help from the environment?
Viruses use their surroundings to replicate themselves. This is very similar to what we are doing. Even we, who consume other animals and plants and inhale oxygen, would lose our lives in a matter of moments if we covered our heads with a plastic bag and separated ourselves from the environment. It can be said that we, like viruses, are parasitic beings on the environment. Similarly, plants need us almost as much as we need them. Plants need sunlight, water, and carbon dioxide (CO2) to produce and grow their own organic matter through photosynthesis. It cannot grow in dry deserts or dark caves, and it is difficult to grow even if there is a lack of CO2. The reason why plants do not run out of this gas is that animals, fungi and various bacteria continuously decompose, digest, and burn organic matter, which is eventually released into the atmosphere as CO2. While burning all of our fossil fuels could have a negative impact on the planet, it is also possible to see this event as a grateful circumstance for plants, as abundant CO2 will allow them to grow more actively. As a result, it can be interpreted that plants, like us, are parasitic on the environment.
From this perspective, the difference between plants, animals, and viruses is just the difference in what they receive from their environment.
Inside the cell, the virus is carefully nurtured in a “perfect womb” that can fill everything. They can live with most things stripped of them, but that is only because the environment around them is very rich. On the other hand, plants make very low demands on the environment around them. It can grow almost anywhere there is light, water and air. Since it survives with very few external factors, the internal mechanism is inevitably complicated. Through their own biochemical mechanisms, plants are able to synthesize everything they need to grow from the air. We humans can be said to exist between viruses and plants. Beyond the universal requirement to eat, we need certain vitamins in our diet, without which we get terrible diseases such as scurvy. Vitamins are compounds that we cannot synthesize ourselves from simple precursors, and we have already lost the biochemical mechanisms that our ancestors had to synthesize them. Without the outside help of this vitamin, we would suffer the same fate as a virus that has no host.
If we all need help from the environment, the only question left is, “How much do we need?”
Dependence on the environment
“Parasites”, which parasitize parts of DNA, such as viruses and “retrotransposons” (Note 16), replicate themselves throughout the genome without leaving a safe host. Both retrotransposons and viruses have the ability to make copies of themselves (self-replication) by using the biological environment around them. In other words, inanimate and animate objects are continuous, and it makes no sense to try to draw a line between them. Most definitions of life focus on living organisms and often ignore environmental parasitism. For example, NASA’s “tentative definition” of life defines life as “a self-sustaining chemical system that can evolve Darwin”. Viruses may not be included in this definition, but it will depend on the interpretation of the vague word “self-sustaining”.
(Note 16) Genes that can move in the genome are called “transposons”. Transposons are divided into “DNA-type transposons” and “retrotransposons (RNA-type transposons)” depending on how they move. In “retrotransposon” the donated DNA is transcribed into RNA (primary copy), which is further copied and DNA is synthesized (secondary copying). Since the copied DNA is inserted, the retrotransposon is characterized by copying and pasting, and the retrotransposon in the donated DNA does not move and remains in its original place. On the other hand, “DNA-type transposons” are characterized by their cut-and-paste ability because they can move with DNA.
The way in which “transposons” move through the genome is shown schematically and in an easy-to-understand manner, for example, on the following site.
In any case, the dependence of life on the environment is not emphasized. The environment, by its very very nature, appears to be outside of life, but it is by no means so, and the two are always inextricably linked.
What would happen if we were separated from the environment to which life is adapted? We humans probably won’t survive and will die. It seems to us that we only have two choices, life and death, but that is not always the case. When disconnected from the resources of the host cell, the virus does not immediately decay and “die”. They can withstand being robbed of this and that by the world. For example, there are 10 times as many viruses in 1 milliliter of seawater as there are bacteria, waiting for that moment. The virus’s ability to resist decay is reminiscent of bacterial “spores” that can remain in a state of asphyxia and live for many years. Spores have no metabolic activity and have the ability to survive in permafrost and even outer space for thousands of years. Not only spores, but also organisms such as seeds and tardigrades can withstand extreme conditions such as complete dryness, radiation 1,000 times the amount that kills humans, tremendous pressure on the ocean floor, and the vacuum of space.
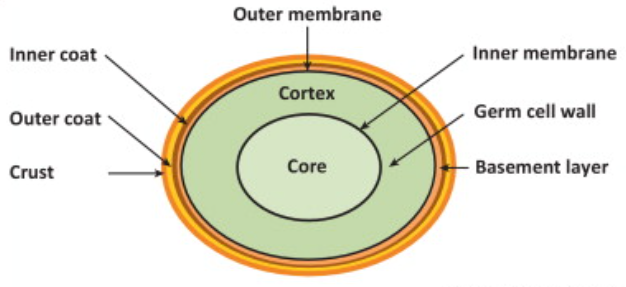
Why don’t viruses, spores, and tardigrades collapse even though they should collapse according to the second law of thermodynamics? They may eventually disintegrate, but in the absence of them, they are surprisingly stable, despite their lack of vital activity.
This tells us an important lesson about the difference between “Life” and “Living.”
- Spores are classified as organisms by many biologists, but they are not strictly alive. Still, they are not dead, as they have the ability to revive and are able to carry out life activities again.
- Viruses also have the ability to self-replicate again when suitable conditions are met. The same applies to tardigrades.
- “Life” is a structure determined by genes and evolution, but “Living” depends on the environment in addition to processes such as growth and reproduction, and this is nothing but the interrelationship between structure and environment.
Energy, Entropy and Structure
According to the “Second Law of Thermodynamics”, “entropy (disorder)” always increases, so at first glance it seems strange that spores and viruses are so stable.
Unlike life, entropy is specifically defined and can be measured, and the units are joules/kelvins/moles. So, let’s crush the spores into pieces and investigate the change in entropy. If you grind it into molecules and measure the change in entropy, you will find that the entropy must be increasing. This is because, when appropriate conditions are met, a wonderfully orderly system that can grow again has become a collection of random, non-functional fragments, it is natural to assume that the entropy is high.
However, according to the precise measurements of bioenergetic researcher Ted Buttley, the entropy remained almost unchanged. This is because entropy involves more than just spores. The environment must also be considered, and there is a certain degree of disorder in it.
Spores are made up of parts that interact with each other tightly fitted. In it, an oily (lipid) film naturally pushes water away by the physical forces acting between the molecules. When oily lipids are mixed with water and shaken, they naturally form a thin “double layer” that forms a biological membrane that covers the aqueous vesicles. This is because it is the most stable state. That is, because of physical attraction and repulsion, oil and water want to act with themselves, not with each other. Proteins behave in much the same way. Those with a large amount of electric charge are soluble in water, and those without an electric charge show “hydrophobicity” that works well with oil. In other words, they hate water. When oily molecules gather and settle, and charged proteins dissolve in water, energy is released. It is the physically stable, low-energy, and “comfortable” state of matter. Energy is released as heat.
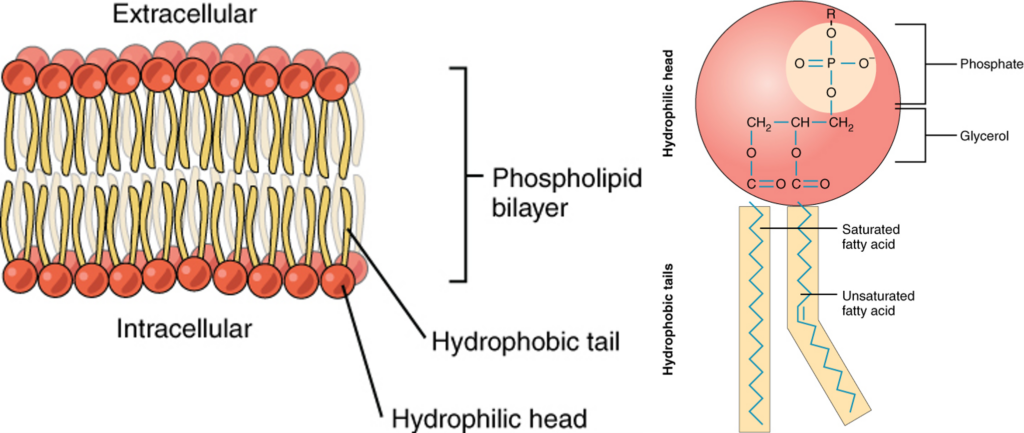
Heat is the motion of molecules , the disorderliness of molecules jostling and colliding with each other, and as a result, it gives rise to entropy. Thus, when oil and water separate and heat is released, entropy increases. Then, from the point of view of overall entropy, if all these physical interactions are taken into account, an ordered membrane surrounds the cell with a higher entropy than a random mixture of unmiscible molecules.
If a physically pleasant state releases energy into the environment as heat, then a physically uncomfortable state does the opposite: it absorbs energy from the environment, lowers the entropy of the environment, and cools the environment. If all of this is taken into account in the case of spores, the overall entropy changes very little.
At the molecular level, the structure of the polymer minimizes energy locally and releases excess energy into the environment as heat, increasing the entropy of the environment. Proteins naturally fold into shapes with the lowest possible energy. Its hydrophobic part is buried away from the surface water. Since electric charges attract and repel each other, positive charges stay in a fixed place in proportion to negative charges, stabilizing the three-dimensional structure of proteins. Thus the proteins are naturally folded into a particular shape, even if they are not necessarily useful.
Perhaps most surprising is the fact that there is little difference in overall entropy between a chaotic soup of disparate amino acids, which are the building blocks of proteins, and a beautifully folded protein. When the folded protein is unfolded, it returns to a state more like a soup of amino acids, increasing entropy. However, doing so also exposes hydrophobic amino acids to water, and this physically uncomfortable state absorbs energy from the outside, reducing the entropy of the environment and cooling the surroundings.
The idea that life exists in a state of low entropy, meaning it is more organized than a disordered soup, is not strictly correct. This is because the order and organization of life is outweighed by the increasing disorder of the environment.
So, what does Erwin Schrödinger mean when he said that “Life sucks up negative entropy from the environment” (Note 17)?
- In those words, he wanted to say that “Life somehow derives order from its surroundings.”
(Note 17) Erwin Schrödinger, “What is Life?”
In fact, even if the soup of amino acids has the same entropy as a tightly folded protein, proteins are less likely to occur and therefore require more energy in two senses. First, soups of amino acids do not naturally combine to form chains. Proteins are chains of amino acids, but amino acids are inherently less reactive. In order for amino acids to bind, living cells must first activate it. Only then can they react and form chains. At this time, about the same amount of energy is released as was used in the initial activation. The energy that proteins release when they fold themselves is lost as heat, increasing the entropy of the environment. So,
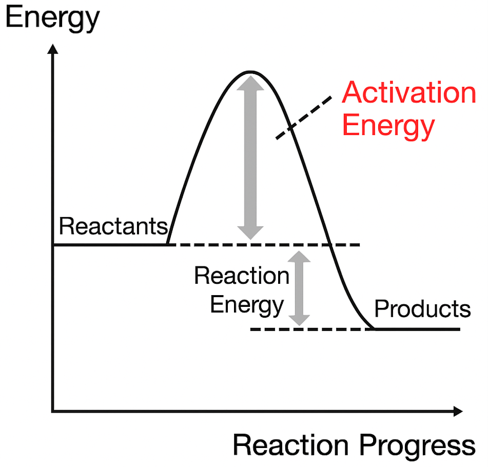
there is also an “energy barrier” (activation energy) between two equally stable states. Just as energy barriers make it difficult to form proteins, there are also barriers to protein degradation. It takes some effort (and digestive enzymes) to break down a protein and turn it back into its constituents.
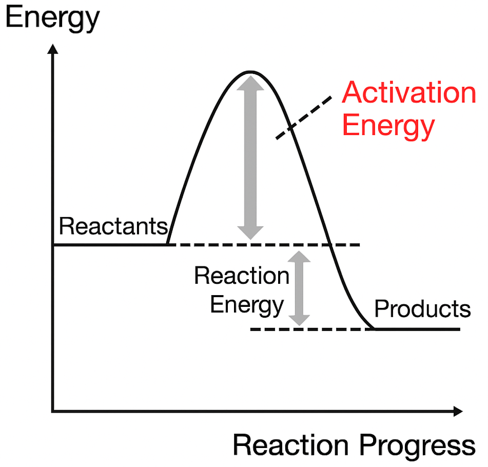
If there are enough reactive components, such large structures will be particularly stable. And the real question is, “Where do these highly responsive components come from?”
Here arises a second problem as well: not only activated amino acids, but even a soup of amino acids cannot be said to exist with certainty under today’s environmental conditions. If left unchecked, it will eventually react with oxygen and revert back to a simpler mixture of gases – carbon dioxide, nitrogen oxides, and water vapor. In other words, it takes energy to make amino acids in the first place, and that energy is released when the amino acids are broken down. Therefore, even if we starve, we can survive for a while by breaking down muscle proteins and using them as fuel. This energy is obtained not by the protein itself, but by the combustion of the amino acids that make it up. Therefore, neither seeds, nor spores, nor viruses are completely stable in today’s oxygen-rich environment. These components slowly react with oxygen over time (oxidized) and eventually undermine their structure and function, making it impossible to resuscitate even under appropriate conditions. The seed dies. However, if you change the surrounding gas and keep oxygen at bay, it will be stable forever. Since living organisms are not in equilibrium due to the oxygen-rich global environment, they are susceptible to oxidation unless they are actively prevented from progressing. Like this,
in a normal environment (in the presence of oxygen), energy is required to make amino acids and nucleotides and other building blocks of living organisms from simple molecules such as carbon dioxide and hydrogen. It also takes energy to connect them into long chains, polymers such as proteins and DNA, but there is almost no change in entropy. That’s what “life” is all about.
- Create new elements, connect them all, grow, and multiply. Growth is also the active transport of substances in and out of the cell. Doing this requires a continuous flow of energy.
- Schrödinger called it “free energy.” The equation he had in mind was symbolic, relating entropy and heat to free energy.
ΔG=ΔH-TΔS
The Greek letter Δ (delta) indicates change, but ΔG is a change in “Gibbs’ free energy”, named after J. Willard Gibbs, the great and lonely American physicist of the 19th century. This energy allows mechanical work such as muscle contraction and intracellular phenomena to be triggered “freely”.
ΔH is a change in heat, which is released into the environment and warms the environment, thus increasing the entropy of the environment. The reaction of releasing heat into the environment cools the system itself. This is because the energy in the system is less than before the reaction. Then, when heat is released from the system to the environment, the ΔH of the system has a negative sign.
T is the temperature, and its importance depends solely on the environment. Releasing a certain amount of heat into a cold environment has a greater impact on that environment than releasing exactly the same amount of heat into a warm environment. This is because the relative input is larger.
Finally, ΔS is the change in the entropy of the system. When the entropy of the system decreases, it becomes a negative sign and increases order, and conversely, when the entropy increases, it becomes positive and the system becomes more disorderly.
Overall, in order for any reaction to occur naturally, the free energy ΔG must be negative. The same can be said of the sum of all the reactions that make up “life”. That is, the reaction occurs spontaneously only when ΔG is negative. In order for this to happen, there is a need for,
- The entropy of the system increases (the disorder of the system increases), or
- Energy is lost as heat from the system,
- Or both.
Therefore, it can be said that local entropy can decrease – the system can increase order – as long as ΔH is more negative (i.e., a large amount of heat is released into the environment). In the end,
- In order to promote growth and reproduction, or “life,” some reaction must constantly release heat into the environment and increase the disorder of the environment.
The stars in the sky pay the price of an orderly existence by releasing enormous amounts of energy into space. In our own case, we pay the price of a sustained existence by releasing heat from the constant reaction of breathing. We are constantly burning food in oxygen and releasing heat into the environment. Its heat loss is not in vain. It is absolutely necessary for the survival of life.
All phenomena that occur in living cells are spontaneous and can proceed on their own if the appropriate starting point is given. ΔG is always negative. Energy follows a downward trajectory all the way. But this means that the starting point must be at a very high point.
If you were to make a protein, the starting point would be a ridiculous assemblage of activated amino acids packed into a small space. These amino acids will release energy as they bind and fold to form proteins, increasing the entropy of the environment.
Activated amino acids can be formed spontaneously if there are enough moderately reactive precursors. In addition, such moderately reactive precursors are also formed spontaneously if given a highly reactive environment. So as it turns out,
- The power of growth comes from the reactivity of the environment, which in our case in the form of food and air, and in the case of plants, in the form of photons, which are constantly flowing through living cells.
- Living cells use this continuous flow of energy to grow and overcome their tendency to degrade.
It is made by an elaborate structure that genes specify to some extent. But whatever such a structure is, it is itself the result of growth and replication, natural selection and evolution.
- None of these causes can occur without a constant flow of energy from somewhere in the environment.
- Living organisms require an enormous amount of energy to survive.
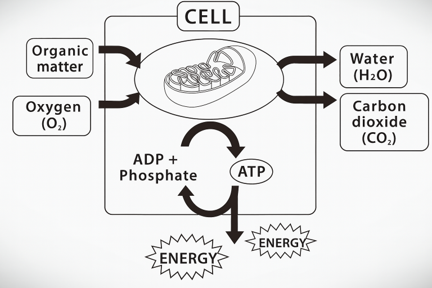
The energy medium used by all living cells, known as the “energy currency,” is a molecule called ATP (adenosine triphosphate). ATP provides energy for certain proteins to change from one stable state to another. In other words, the protein changes from one stable three-dimensional structure to another. However, after this change, the protein stops moving again. In order to restore the structure, ATP is required again.
In the cell there is a large number of proteins, which are driven by ATP. Therefore, it can be said that one cell consumes about 10 million ATP molecules per second. The number is surprisingly high. There are about 40 trillion cells in the human body, and the total amount of ATP they consume and regenerate is about 60 to 100 kilograms per day, which is about the same amount as our own body weight. However, in reality, the ATP present in our body is about 60 grams, and this fact suggests that each ATP molecule is replenished once or twice a minute.
The mechanism of its replenishment is as follows.
- When ATP is decomposed, it releases free energy that causes changes in the three-dimensional structure. At the same time, it also releases enough heat to keep ΔG negative. ATP is usually divided into two parts: “ADP (adenosine diphosphate)” and “inorganic phosphate (PO43-)”. This inorganic phosphoric acid is the same as that used in fertilizer and is usually labeled Pi.
- It also takes energy to form ATP again from ADP and Pi. The energy obtained by respiration (the energy released by the reaction between food and oxygen) is used to regenerate ATP from ADP and Pi.
- Here is the complete picture. This circulation of energy is the whole picture of the energy metabolism cycle of life.
ADP + Pi + Energy ⇋ ATP
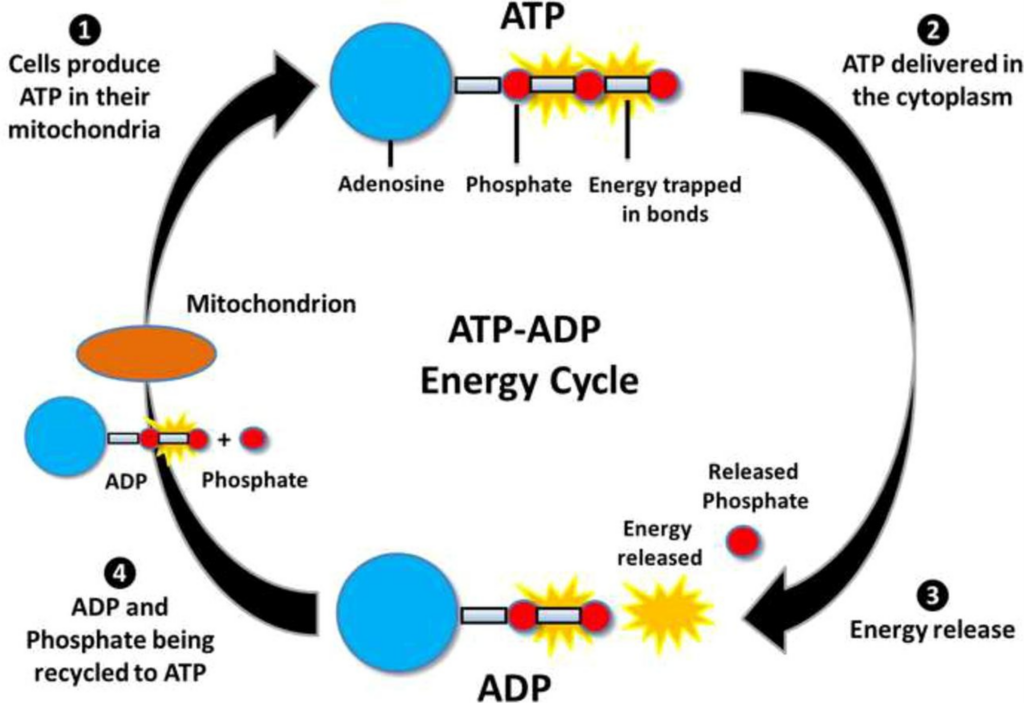
Our human consumption of ATP is relatively low. For example, bacteria such as “Escherichia coli”, which divides every 20 minutes, consume about 50 billion ATPs per cell division as energy for growth. This is about 50~100 times the weight of a single cell, which is almost 4 times the rate of our ATP synthesis. If we convert this energy consumption in watts, we can see that the energy consumption of life is implausible as follows.
- We humans consume about 2 milliwatts of energy per gram of body weight. Thus, the average human weighing 65 kilograms consumes about 130 watts of energy, which is just a little more than a standard 100-watt light bulb.
- This number may not seem large, but per gram it is 10,000 times the energy consumed by the sun.
- In fact, at any given moment, only a small part of the Sun is undergoing nuclear fusion.
- The use of life energy is more like a rocket on a launch pad than a candle.
Therefore, from a theoretical point of view, life is not enigmatic and does not violate the laws of nature. The amount of energy consumed by living cells per second is enormous, but the amount of solar energy that falls on the earth is significantly higher. The sun’s energy is less powerful per gram, but its size makes up for it. If part of this energy drives biochemical mechanisms, then it is conceivable that life can operate in almost any way. There seems to be no fundamental constraint on the use of energy.
However, it is a surprising fact that life on Earth is extremely constrained from the perspective of energy utilization.
Specifically, there are two surprising characteristics regarding the use of energy in life.
- The first is the fact that the energy of all cells is derived from a type of chemical reaction called a “redox reaction.” This reaction is “a phenomenon in which electrons are transferred between molecules”.
- “Redox” is a term that refers to “reduction and oxidation,” which refers to the transfer of one or more electrons from a donor to a receptor. It is thought that the body donated with electrons is oxidized.
This phenomenon occurs when iron reacts with oxygen, and iron donates electrons to oxygen and oxidizes itself. The substance that received the electron – in this case, oxygen – is reduced. For example, in respiration and combustion, oxygen (O2) is reduced to water (H2O). This is because the oxygen atom acquires two electrons and two protons to maintain the balance of charges.
This reaction releases energy as heat, and the energy of the system itself that causes the reaction decreases. In particular, in the reaction between iron and food and oxygen, a large amount of energy is released like combustion.
by respiration
Respiration stores some of the energy released in this reaction in the form of ATP, at least for a short period of time until ATP is broken down again. During decomposition, the energy remaining in the ADP-Pi bond of ATP is released as heat. After all, respiration and combustion are equivalent, and the slightest delay in the process is recognized as “life.”
A series of redox reactions is ultimately the movement of electrons along a chain of carriers, similar to an electric current flowing through a conductor. In breathing, such electron transfer is carried out.
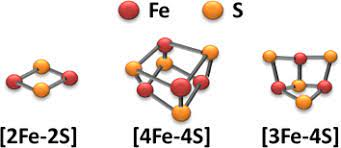
Electrons stolen from food are not passed directly to oxygen, but rather to “stepping stones” (usually part of ionized iron atoms (Fe3+) found in respiratory proteins, often in small inorganic crystals called “iron-sulfur clusters”) moving on. From there, the electrons jump to clusters that are very similar but have a slightly higher “need” for electrons. When an electron is attracted to the next cluster and crosses, the cluster is first reduced (Fe3+ becomes Fe2+ because it receives electrons) and then oxidized (it loses electrons and returns to Fe3+). After repeating this process about 15 times or more, the electrons eventually reach oxygen.
At first glance, the means of growth, such as photosynthesis in plants and respiration in animals, seem to have little in common, but they are actually basically the same in that they involve the movement of electrons following such a “respiratory chain.”
Life can function using any imaginable type of energy, including thermal energy, mechanical energy, radioactivity, electrical discharge, and ultraviolet radiation. Surprisingly, however, all life forms acquire energy in a “redox reaction” through an almost similar respiratory chain.
Why?
The second surprising aspect of the energy of life is the specific mechanism by which energy is stored in the binding of ATP.
- Life does not use simple chemical reactions, but promotes the production of ATP by means of a “proton gradient” (Note 18) that separates thin membranes.
- Today, the molecular-level mechanisms by which “proton gradients” occur are well understood, and we also know that proton gradients are ubiquitous in life on Earth. The “power of protons” is as essential to life as DNA, the universal genetic code.
(Note 18) The “proton gradient” is a gradient of electrochemical potential, and refers to the “protons (hydrogen ions)” that move across the membrane. The gradient consists of two components: the “chemical gradient” (the difference in solute concentration across the membrane) and the “electrical gradient” (the difference in electric charge across the membrane). When the proton concentration on both sides of the permeable membrane is uneven, the proton moves across the membrane by simple diffusion from the high concentration side to the low concentration side. Since the proton has an electric charge, an electric potential is also formed between the membranes. If the charge is unevenly distributed across the membrane, the potential difference creates a force that drives the diffusion of the proton until the charge is equal on both sides of the membrane.
However, little is still understood about how this mechanism responsible for the energy generation of life forms evolved.
Whatever the reason, life on Earth seems to be using only a surprisingly limited and peculiar subset of possible energy-producing mechanisms – the “redox reaction” and the “proton gradient”. Is this an indication of a historical coincidence, or did these methods become widespread because they were superior to any other, or is the final question whether they are the only possible methods? These questions still await answers.
ATP synthase in the mitochondria
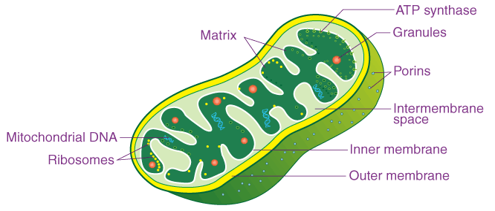
Therefore, we start by observing the part of cellular respiration deep in the “mitochondria”, the “thermodynamic center of the cell”, where energy generation of living organisms is carried out.
Here, the protons are stripped of the molecules of what you eat and left behind, and passed on to “Complex I”, the first and largest component of the massive “Respiratory Chain Complex”(Note 19).
(Note 19)The “respiratory chain complex” is a giant protein with a molecular weight of about 100,000 to 1,000,000 that exists in membranes (inner mitochondrial membranes, thylakoid membranes, and prokaryotic cell membranes) found in most organisms that perform cellular respiration regardless of aerobic or anaerobic respiration. It consists of “respiratory chain complex I, II, III, IV”, and ATP synthase is also called “respiratory chain complex V”.
This huge complex I has a complex structure of 45 proteins, each of which is composed of hundreds of amino acids. If ATP is about the size of a human being, then Complex I is about the size of a skyscraper. Moreover, these are not just giant structures, but huge dynamic machines that work like steam engines.
In this complex I, electrons are carried separately from the proton. It is taken in from one side and discharged from the depths of the other membrane. After that, the electron goes through two more huge protein complexes. The sum of these is called the “respiratory chain”. Each complex contains multiple “redox centers” (about nine in Complex I) and has the function of temporarily holding electrons. Electrons leap from one of these “centers” to the next. The fact that these “centers” are evenly spaced suggests that electrons exhibit a “quantum tunneling effect.” The electron’s eye can only see the next redox center, unless it is far away.
Here, the distance is measured in units of angstroms (Å) (1 Å is approximately the size of one hydrogen atom). Each redox center is within 14 Å of the following spacing. Then, as long as the affinity for the electron is slightly higher than the previous redox center, the electron will leap along the continuous path of the redox center. This is similar to the image of crossing a river by following a series of stepping stones at appropriate intervals.
(Note 20)Nicotinamide adenine dinucleotide is an electron carrier and energy carrier used by all eukaryotes and many archaea and eubacteria. This substance functions as a coenzyme for various dehydrogenases and can take on two states: oxidized (abbreviation NAD+) and reduced form (also written as NADH or NADH2). It undergoes two-electron reduction, but does not produce an intermediate form. These are used in general metabolism such as glycolysis, TCA cycles, and respiratory systems that synthesize ATP.
Electrons cross three huge respiratory chain complexes at once. The electrons are attracted forward by the strong force of oxygen. This differs from remote action in that it is a matter of probability that electrons are more attached to oxygen than elsewhere. After all, this phenomenon is separated by proteins and lipids, which are conduits for the flow of electrons from food to oxygen. This current moves everything in that place. Electrons leap along the path they need to take, and they are only interested in the route they take to reach oxygen, and they don’t see the machines around them.
However, there are a large number of interlocking switches in these huge protein complexes. When electrons are present in the redox center, neighboring proteins take on a certain structure. Its structure changes slightly as the electron moves to the next redox center, the location of the negative charge shifts, the positive charge follows, and the entire network of weak bonds is readjusted, transforming the massive structure into a new three-dimensional structure in an instant. When a small change occurs in one part of a protein, a cavernous channel opens somewhere else. Then, a new electron arrives, and the machine returns to its previous state.
This process is repeated dozens of times per second. Much is now understood about the structure of these respiratory chain complexes with a resolution of only a few angstroms at the almost atomic level. We know how protons bind to water molecules that are fixed in place by the charge of the protein. We also know how these water molecules move when the channel itself changes its structure.
In addition, it is well understood how protons can quickly pass through gaps that dynamically open and close and are exchanged between water molecules. It closes as soon as the peloton passes, preventing a turning back. This enormous and elaborate mechanism of motion plays one role in transporting protons from one side of the inner mitochondrial membrane to the other.
The first complex of the respiratory chain sends four protons across the membrane for every two electrons that pass. The two electrons then enter the second complex (which is technically complex III, which has a path independent of complex I), which sends four more protons across the membrane. Finally, in the final respiratory chain complex, the electron meets oxygen, but before that, two more protons cross the membrane. As a result, for every two electrons extracted from food, a total of 10 protons are transported across the membrane.
A little less than half of the energy released by the flow of electrons towards oxygen is stored in the proton gradient. All of its power, all of its cleverness, and all of its massive protein structure are directed exclusively to pumping out protons beyond the inner membrane of the mitochondria.
In one mitochondria, there are tens of thousands of respiratory chain complexes. A single cell contains hundreds to thousands of mitochondria. There are at least 1,000 trillion mitochondria in the 40 trillion cells in your body, and the total surface area of these intricate membranes is about 14,000 square meters. That’s the size of four football fields.
The main role of mitochondria is the extraction of protons, and as a whole, more than 1021 protons are extracted per second. This number is close to the number of stars in the known universe.
But this is only half the work of mitochondria. The other half is to harness that energy to produce ATP.
The membrane of the mitochondrion is almost impermeable to protons, and any dynamic channels that allow proton passage close immediately after the protons pass through. Protons are very small (nothing more than hydrogen nuclei) and because of their size, it is not easy to block them. Since the proton passes through the water molecules instantly, the membrane must be completely sealed to prevent water from entering. In addition, the proton has a positive charge.
When the proton is pumped out beyond the sealed membrane, two things are accomplished.
- First, there is a difference in proton concentration (hydrogen ion concentration) on both sides of the membrane.
- Second, it also creates a difference in charge, where the outside is more positive than the inside. Then, an electrochemical potential of about 150 ~ 200 millivolts is generated across the membrane. Since the film is very thin (about 6 nanometers thick), this difference in charge is enormous over short distances. The strength of the electric field perceived near the membrane, is 30 million V/m, which is comparable to a lightning bolt.
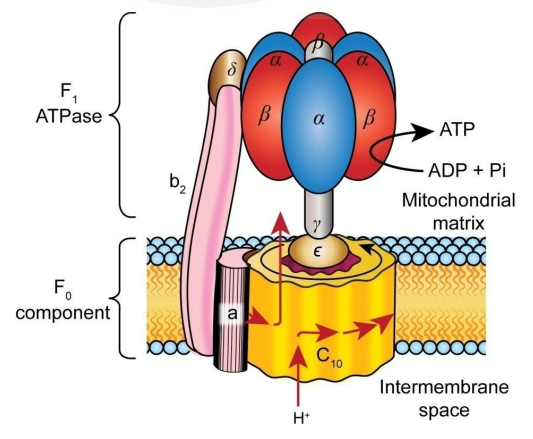
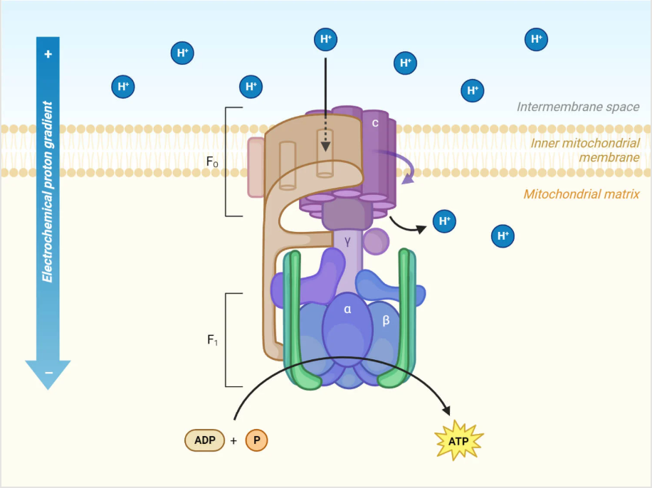
This large electrochemical potential is called the “proton driving force” and drives the “ATP synthase” (Figs. 34~36), which can be said to be the most magnificent protein nanomachine. Drive refers to motion, and ATP synthase is in fact a rotary motor (the F₀ motor), in which the flow of protons turns a crankshaft, which in turn rotates the catalyst site head (catalyst head, F1 motor) . This mechanical force proceeds with the synthesis of ATP. Metaphorically, this protein acts as a turbine in hydropower, whereby the proton, trapped in a reservoir beyond the membrane barrier, rushes into the turbine like flowing water, turning a rotating motor.
Its mechanism of action is not yet fully understood.
- How individual protons are connected to the C-ring (C-subunit, the rotor that makes up the motor part) in the membrane,
- How this rotating ring twists the crankshaft, resulting in a structural change in the catalyst head,
- How the void that opens and closes with this head grabs the ADP and Pi and mechanically binds them together to produce new ATP.
The understanding of above things is incomplete. This is a magical device that can be said to be the highest level of precision nanoengineering (Fig. 36), and the more you learn about it, the more you will be amazed.
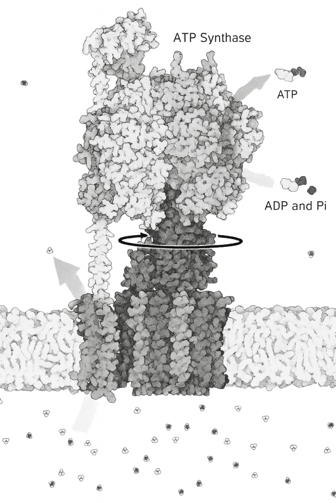
In ATP synthases, every time 10 protons pass through, the rotating head goes all around and three newly synthesized ATP molecules are released as substrates (substances that catalyze chemical reactions by enzymes). The head can rotate at a speed of more than 100 times per second.
ATP is called the universal “energy currency” of life, but it is also called “ATP synthase” and “proton driving force”is also universally preserved throughout the evolutionary history of life as a whole. These are as universal as the genetic code. Therefore,
- “ATP synthase”, like the “DNA double helix”, can be regarded as one of the defining symbols of life.
The concept of the “proton driving force” was created by Peter Mitchell, one of the scientists who revolutionized the field of bioenergetics in the 20th century. He published the “chemiosmotic theory” in 1961, and “chemiosmotic” was Mitchell’s term for the movement of protons across a membrane. The word “osmotic” in this case means “pushing” and is different from the commonly understood usage of “water passing through the semipermeable membrane”.
In other words, “breathing” pushes protons beyond the thin membrane against the concentration gradient, and Mitchell used the expression chemiosmotic (chemistry + pushing). In other words, respiration is not a chemical reaction by a reaction medium, as was thought until the middle of the 20th century.
- The mechanism that links the flow of electrons to ATP synthesis is actually a “proton gradient – proton driving force” that occurs between semipermeable membranes.
Basically, all life uses a “redox reaction” to create a proton gradient across the membrane. Why on earth does life depend on such mechanisms?
That is,
- Why do all living cells use the “redox reaction” as a source of free energy?
Thinking even more deeply, one arrives at the question,
- Why “electron”? Why “proton”?
Therefore, I would like to discuss these two basic questions.
Life is electrons after all
So why does life use redox reactions?
The basis of life as we know it is carbon, specifically “carbon in a partially reduced form”. Elements such as nitrogen and phosphorus are also required in relatively small amounts, but with extreme approximation, it can be said that the “chemical formula of life” is “CH2O”. Considering carbon dioxide as a starting point, life requires the transfer of electrons and protons from hydrogen (H2) to CO2. In theory, the source of the electron can be anything. It is also possible to take electrons from water (H2O), hydrogen sulfide (H2S), and even ferrous iron (Fe2+). What you mean,
- it is crucial that electrons are transferred to CO₂, and all of these transfers are redox reactions. By the way, the expression “partially reduced” refers to the fact that CO2 is not completely reduced to methane (CH4).
What if life could use elements other than carbon? Other elements, such as silicon, may also be considered. However, carbon has many advantages. that is
- A single carbon atom can form four strong bonds, which are much stronger than those formed by chemically close silicon.
- This bond allows the creation of a wide variety of long-chain molecules, especially proteins, lipids, sugars, and DNA.
- With silicon, it is difficult to achieve such a variety of chemical reactions. In addition, there is no oxide of gas comparable to carbon dioxide in silicon.
If you think of CO2 as Lego bricks,
- It is taken out of the air and can add one carbon to other molecules at a time.
- Silicon oxide, on the other hand, is like trying to build with sand. Elements such as silicon may be suitable for use by relatively advanced intelligent life such as humans, but the possibility of using silicon to form life from scratch is difficult to see.
Also, as a matter of probability and predictability, the possibility seems overwhelmingly low. In addition to being much better, carbon is also much more abundant throughout the universe. Therefore, roughly speaking, life should be carbon-based.
So, let’s look at the problem from the other side. Specifically, we will explore why the “redox reaction” is useful.
When thinking about “breathing”, we should take into account not only humans, but also other life forms. We take electrons from food and carry them to oxygen through the respiratory chain.
The important point here is that it is also possible to replace both the source of electrons and the place of breathing with different ones.
When food is burned in oxygen, the efficiency of energy collection is maximized, but the underlying principle is very diverse and flexible. In fact, it is not always necessary to consume organic matter. “Hydrogen gas”, “hydrogen sulfide”, and “ferrous iron” are all electron donors, as described above, and if there is a sufficiently strong oxidizing agent on the other side, it is possible to pass its own electrons into the respiratory chain. Therefore, bacteria can “eat” rocks, minerals and gases using essentially the same protein apparatus that we use for breathing.
The presence of oxygen is also not mandatory. Many other oxidizing agents can perform almost the same task. “Nitrates”, “nitrites”, “sulfates”, “sulfites”, and the list goes on and on. All of these oxidizing agents (so called because they exhibit a reactivity similar to oxygen) have the ability to absorb electrons from sources such as food. Bacteria can not only “eat” rocks, but also “breathe” (Note 21).
(Note 21) When an organism acquires energy through respiration, electrons released along with the metabolism of organic matter are passed to electron acceptors such as oxygen. At this time, energy equivalent to the difference in redox potential between the electron donor (such as NADH(Note 22) produced by metabolism) and the electron acceptor is released, and organisms mainly use this energy to synthesize ATP. Th e type of respiration that uses substances other than oxygen as the electron acceptor is called “anaerobic respiration”, and bacteria or archaea that live in oxygen-free environments (“anaerobic bacteria”) use substances such as nitric acid, sulfuric acid, and carbon dioxide as electron acceptors. Some anaerobic bacteria use solid metal compounds such as iron oxide as electron acceptors to breathe (“iron-reducing bacteria”), and these bacteria have pathways for transmitting electrons to solids that exist outside the cell (extracellular electron transfer pathways).
(Note 22) “NADH (nicotinamide adenine dinucleotide)” is an electron carrier used in all eukaryotes and many archaea and eubacteria. It acts as a coenzyme for various dehydrogenases and can take on two states: oxidized and reduced. It undergoes two-electron reduction, but does not produce an intermediate form. The abbreviation NAD⁺ is more commonly used in papers and oral speeches.
This diversity in terms of electron donors and receptor utilization is aided by the slow response of many of them. All biochemical mechanisms arise spontaneously and must be driven by a highly reactive environment. However, if the environment is too reactive, the reaction will proceed quickly, and there will be no free energy left to drive life activity.
For example, the atmosphere will not be filled with fluorine gases. This is because it immediately reacts to anything and disappears. However, many substances react so slowly that they accumulate at levels well beyond their natural thermodynamic equilibrium. If given the opportunity, oxygen would react violently with organic matter and burn everything on the planet, but this violent tendency has been tempered by the fortunate chemistry that has kept the Earth stable for so many years. Gases such as methane and hydrogen react more violently with oxygen than organic matter, but there are kinetic barriers to these reactions, so all of these gases can coexist in the air for years in a state of “dynamic imbalance” (Note 23). The same is true for many other substances, from hydrogen sulfide to nitrate. They can be forced to react, at which time a large amount of energy is released and available to living cells, but without the right catalyst, much does not happen.
(Note 23) The reaction of 2H2 + O2 → 2H2O is an exothermic reaction, and since the free energy is also reduced, it is a spontaneous reaction. However, the reaction does not occur just by mixing H2 and O2, and it does not occur unless the temperature is quite high. This is considered as follows. There is an “energy barrier” that causes a reaction, and molecules are usually unable to overcome this energy barrier. However, as the temperature rises, some molecules begin to emerge with enough energy to overcome the barrier, and the reaction proceeds. This energy barrier is called “activation energy”. In the case of nA + mB→C, the reaction rate v can generally be written as follows.
v= k[A]n[B]m
Here, k is referred to as “reaction rate constant”. There is a following important relationship between the reaction rate constant k, the activation energy Ea and the temperature T,
k=αe-Ea/RT
Life uses these “kinetic barriers” to increase entropy faster. In this respect, some even define life as an “entropy generator”. Anyway,
- Life exists precisely because of the presence of kinetic barriers, and it excels at overcoming them.
- Without the escape route of high reactivity, which is held back beyond those kinetic barriers, it is doubtful whether life could have existed at all.
Moreover,
- The fact that many electron donors and acceptors are water-soluble and chemically stable, and can easily enter and exit cells, means that a highly reactive environment required by thermodynamics can reliably be brought into the cell and to its crucial membranes.
- This makes the “redox reaction” much easier to work with as a form of biologically useful energy flow than thermal energy, mechanical energy, ultraviolet radiation, or lightning.
Surprisingly, respiration is also the basis of photosynthesis. There are several types of photosynthesis, in each of which a pigment (usually chlorophyll) that absorbs the energy of sunlight as photons excites electrons and sends them to a chain of redox centers, where they reach acceptors (in this case, carbon dioxide).
The pigment that has lost an electron replenishes it from a nearby donor—be it water, hydrogen sulfide, or ferrous iron. As in the case of respiration, the type of electron donor does not matter in principle. “Oxygen-free” photosynthesis uses hydrogen sulfide and iron as electron donors, leaving sulfur and rusty iron precipitates as waste products. On the other hand, “oxygen-evolving” photosynthesis uses a more difficult donor – water – and excretes oxygen as a waste product.
What is important, however, is the fact that all these various types of photosynthesis
- clearly originate from respiration.
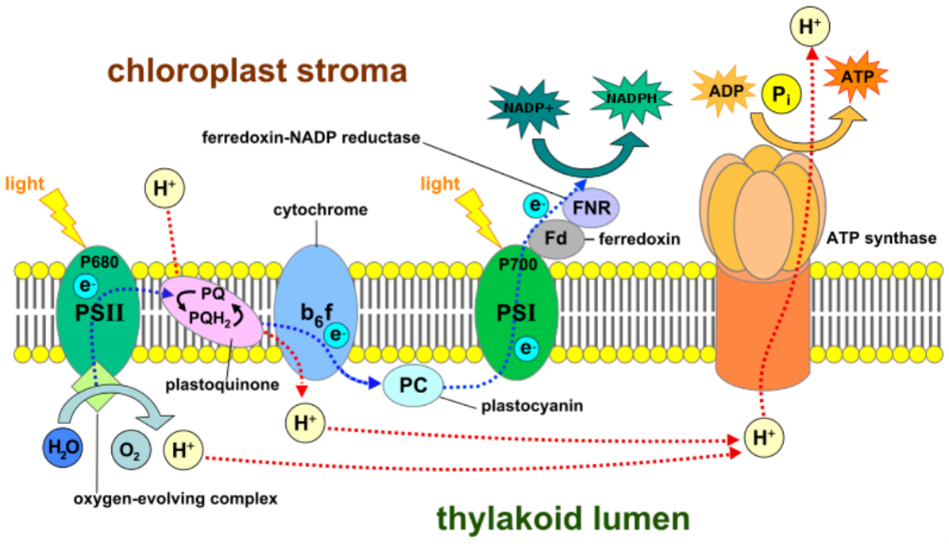
As shown in Fig. 37, exactly the same respiratory system protein, the same type of redox center, the same proton gradient across the membrane, and the same ATP synthase are used.
- The only difference is the change in pigment, and “chlorophyll” is closely related to the pigment “heme” (Note 24), which was used in many ancient respiratory proteins. The use of solar energy changed the world, but from a molecular point of view, it only allowed electrons to flow faster in the respiratory chain.
(Note 24) When “chloroplasts” receive sunlight, they use water taken in from the roots and carbon dioxide in the air to produce starch and release oxygen. At this time, “chlorophyll” plays a role in absorbing the energy of sunlight, and this kind of function is called “photosynthesis”.
In this context, “heme” fulfills various functions, such as electron transfer reactions, redox reactions, and the elimination of reactive oxygen species, by binding to apoproteins within chloroplasts. In addition, “heme” is a “porphyrin complex” (a general term for colored metal complexes in which porphyrin or its derivatives act as ligands) with iron as its central metal. Porphyrin itself is a large, planar macrocyclic molecule that binds to a metal atom via four central nitrogen atoms. Utilizing the property of iron to alternate between the ferrous (Fe²⁺) and ferric (Fe³⁺) states through redox reactions, as well as the ability of ferrous iron to bind molecular oxygen, heme serves roles in electron transport, oxygen transport, and as the active center of various enzymes.
Consequently,
- The great advantage of respiration is tremendous versatility. Nearly any redox pair (any electron donor and electron acceptor pair) can be used to channel electrons into the respiratory chain.
The protein that extracts electrons from ammonium ions is slightly different from the one that extracts electrons from hydrogen sulfide, but their primary function is the same, making them closely related variations. Also, at the opposite end of the respiratory chain, proteins that pass electrons to nitrates and nitrites are different from proteins that pass electrons to oxygen, but they are all related. They are similar enough to each other that they can be used interchangeably.
These proteins can be connected to a common operating system, allowing them to be adapted to any environment. Not only are they interchangeable in principle, but in practice they are freely exchanged in various places. In recent decades, it has become clear that horizontal gene transfer (passing small cassettes of genes between cells like coins) is a frequent occurrence in bacteria and archaea. The genes encoding respiratory proteins are most often exchanged by horizontal transfer. Collectively, biochemist Wolfgang Nitschke calls them “redox protein building kits.”
- Considering all these factors, it can be said that the redox reaction is important for all life in the universe.
- Other forms of energy sources are conceivable, but given the need for redox reactions to reduce carbon and the many benefits of respiration, it is not surprising that life on Earth uses redox reactions as an energy source.
However, the essential mechanism of respiration, the “proton gradient” across the membrane, is a completely different matter.
- The fact that respiratory proteins are distributed in different locations by horizontal gene transfer and mixed together to function in any environment stems from the fact that there is a common operating system.
- However, it will be difficult to predict the “nature of life” that may exist on Earth or elsewhere in the universe unless a clear reason is known why life makes use of protons and why proton gradients should be involved in redox reactions.
Life is a proton after all
The evolution of “chemical osmotic conjugation” (Note 25) is still shrouded in mystery. The fact that all life uses chemoosmolality suggests that chemosmotic conjugation may have occurred very early in evolution. If it had emerged later, it would be difficult to explain how and why it became universal, or why the proton gradient completely replaced everything else. It is extremely rare to find a mechanism that is so universally seen.
(Note 25) Refers to the mechanism by which “chemical synthesis of ATP” and “membrane permeation (osmosis) of protons using the energy of the electron transport chain” combine to perform energy conversion in a cell.
Here, “chemical osmosis” means that ions move across the semipermeable membrane according to an electrochemical gradient. In the process of cellular respiration and photosynthesis, “adenosine triphosphate (ATP)” is produced by the movement of hydrogen ions (H+) across the membrane. Hydrogen ions (protons) diffuse from a region of high concentration to a region of low concentration, and the electrochemical gradient of the protons across the membrane is used for the synthesis of ATP. This process is associated with “osmosis” in which water diffuses across the membrane, hence it is called as “chemical osmosis”. ATP synthases are enzymes that use chemical osmosis to synthesize ATP. ATP synthases allow the passage of protons across the membrane and use the free energy difference to phosphorylate adenosine diphosphate (ADP) to synthesize ATP. ATP production by chemosmosis also occurs in most bacteria and archaea, along with mitochondria and chloroplasts.
As an example, let’s take “bacteria” and “archaea”. All life forms share a “genetic code”, and some basic information processing is also universally preserved. For example, in all living cells, “DNA” is transcribed into “RNA”, which is then physically translated into proteins by nanomachines called “ribosomes”. However, the difference between “bacteria” and “archaea” (Note 26), which are the two major domains of “prokaryotes” (single-celled organisms that do not have a nucleus that encloses DNA in the cell), is very large.
(Note 26) It is one of the major lineages of living organisms. Together with “bacteria” and “eucaryota”, it divides the entire biological kingdom into three. “Archae” are similar in morphology and name to bacteria, but belong to a different lineage from bacteria, and their ecological mechanisms and genes are completely different. “Halobacteria”, “methane bacteria” and “thermophilic bacteria” are well known.
In terms of physical appearance, it is difficult to distinguish between bacteria and archaea, but in many biochemical and genetic characteristics, these two domains are quite different. For example, the specific mechanism of DNA replication, which is considered as fundamental to life as the genetic code, is found to be quite different between bacteria and archaea, including almost all the enzymes that are needed. Moreover, the “cell wall” (a structure found in the cells of plants, fungi, and bacteria but absent in animal cells, and which constitutes part of the extracellular matrix as it lies outside the cell membrane) also differs completely in chemical composition between bacteria and archaea, and their biochemical pathways for fermentation are likewise distinct. Even the “cell membrane”, which is essential for chemiosmotic conjugation, is biochemically different between bacteria and archaea. In other words, the barriers that separate the inside and outside of the cell and the replication of genetic material are not fundamentally conserved. Despite the fact that nothing should be more important for the life of a cell than these.
These are very fundamental differences that raise serious questions about the common ancestor of both bacterial and archaeal groups. If common traits are inherited from a common ancestor, but different traits arise independently of the two lineages, what kind of cells are considered to have been ancestors?
DNA transcription, ribosomal translation, ATP synthase, and partial amino acid biosynthesis were present, but beyond that, there is little in common between the two groups. Among these many differences, only “chemical osmotic conjugation” is universal.
On the other hand, however, there are some difficult problems with the idea that chemiosmotic conjugation arose very early in evolution.
One is the sheer complexity of the mechanism. We have already gained experience with large respiratory chain complexes and ATP synthases (tremendous molecular machines with pistons and rotating motors), but it is questionable whether these could have been created at the earliest stages of evolution, before the advent of DNA replication.
The second problem is the membrane itself, which has achieved a frightening complexity at an early stage. In modern cells, chemiosmotic conjugates work only if the membrane is almost impervious to protons. However, all experiments with what are considered to be early membranes indicate that they were well permeable to protons. In fact, it is extremely difficult to keep the proton impervious to it. The problem is that chemiosmotic conjugates appear to be useless until a number of complex proteins are embedded in a membrane that is impervious to protons. It is only useful when the protein is embedded in the membrane. If so, how did these parts evolve first?
A number of major questions have been raised about the “evolution of life” on Earth. I’ll leave the details for another time, and the questions are as follows:
- Why did life come into existence so quickly?
- Why morphological complexity has stagnated for billions of years
- Why do complex eukaryotic cells only occur once in 4 billion years?
- Why do all eukaryotes have so many inexplicable traits not found in bacteria and archaea, from sexual reproduction and two sexes to aging?
Here are two more questions that are equally disturbing to these questions.
- Why does all life conserve energy in the form of a proton gradient through a membrane?
- How and when did this strange but basic process evolve?
are added. In fact, these two questions are interrelated as a whole.
Therefore, in the next chapter, while answering these questions,
- A naturally occurring proton gradient gave birth to life in a very unique but almost certainly universal environment in the universe. All they needed for the environment were rocks, water, and CO2.
In addition
- Chemioosmotic conjugates have constrained the evolution of life on Earth to the complexity of bacteria and archaea over billions of years.
- The mere occurrence of one bacterium somehow entering another allowed these bacteria to overcome the intense constraints that were constantly being placed on them. The eukaryotes born from this endosymbiosis have an order of magnitude larger genome, which became the raw material for their morphological complexity.
- The close relationship between the host cell and its internal symbionts (which become mitochondria) was behind many of the puzzling properties common to eukaryotes.
Let’s try to figure them out.
Evolution is thought to tend to unfold along a similar path, guided by similar constraints, anywhere in the universe.
Figures and Tables
Fig. 25 Spores of Bacillus subtilis
Kai Reineke, Alexander Mathys, Volker Heinz and Dietrich Knorr, “Mechanisms of endospore inactivation under high pressure” Trends in Microbiology 21 (2013) 296(https://www.sciencedirect.com/science/article/abs/pii/S0966842X13000383)
Fig. 26 Example of lipid bilayer and phospholipid molecule
Lumen Learning, Module 4: The Cellular Level of Organization, The Cell Membrane(https://courses.lumenlearning.com/suny-ap1/chapter/the-cell-membrane/)
Fig. 27 Energy barrier (activation energy)
English version of the figure from the following reference, redrawn by ChatGPT
What are exam notes, chemistry, and reaction kinetics? Activation energy is also explained in detail!
(http://www.jukenmemo.com/chemistry/theory/reaction-rate/)
Fig. 28 ATP-ADP energy cycle
Ralf Jäger, Martin Purpura, John A. Rathmacher, John C. Fuller Jr., Lisa M. Pitchford, Fabricio E. Rossi, Chad M. Kerksick, Health and ergogenic potential of oral adenosine-5′-triphosphate (ATP) supplementation, Journal of Functional Foods 78, March 2021, 104357(https://www.sciencedirect.com/science/article/pii/S1756464621000062?via%3Dihub)
Fig. 29 Reaction in which oxygen oxidizes iron
Slideplayer, Slides of Electrochemistry, Chemical reactions and Electricity(https://slideplayer.com/slide/16456765/96/images/3/Oxidation+of+Iron+Electron+transfer+of+iron-+Electron+transfer+to+oxygen.+Fe+%EF%81%A7+Fe3%2B+%2B+3e-+1%2F2+O2+%2B+2e-+%EF%82%AE+O2-.jpg)
Fig. 30 Synthesis of ATP and release of energy by respiration
English version of the figure from the following reference, redrawn by ChatGPT
NHK High School Course, Fundamentals of Biology, 6th Lecture Breathing(https://www.nhk.or.jp/kokokoza/tv/seibutsukiso/archive/resume006.html)
Fig. 31 Typical iron-sulfur (Fe-S) cluster
Department of Molecular Biology, Faculty of Science, Saitama University, Laboratory of Molecular Control, Research “Iron-sulfur clusters and iron-sulfur proteins”
(http://park.saitama-u.ac.jp/~tougyo/yan_jiu_nei_rong.html)
Fig. 32 Structure of mitochondria
learning programs of BYJU’S, ed-tech company in India
(https://byjus.com/biology/mitochondria/)
Fig. 33 Mitochondrial electron transport system and ATP synthesis
Wikipedia, the free encyclopedia ” Oxidative phosphorylation”
(https://en.wikipedia.org/wiki/Oxidative_phosphorylation)
Fig. 34 Mitochondria and ATP synthase
Sanju Tamang“ATP Synthase: Structure, Mechanism, Significances” Biochemistry, September 13, 2023, Microbe Notes
(https://microbenotes.com/atp-synthase/)
Fig. 35 Structure of ATP synthase
Unit 2 ELECTRON TRANSPORT SYSTEM IN PLANTS, eGyanKosh- a National Digital Repository(https://egyankosh.ac.in/bitstream/123456789/104754/3/Unit-2.pdf)
Fig. 36 ATP synthase
English version of the figure from the following reference, redrawn by ChatGPT
THE VITAL QUESTION “Why Is Life the Way It Is?” Nick Lane, translated by Takao Saito, Misuzu Shobo Co., Ltd., 1st printing on September 13, 2016, 10th printing on March 9, 2018 Figure 10 “Structure of ATP synthase”, p. 85
Fig. 37 Flow of photosynthetic light reaction
Melissa Ha, Maria Morrow,& Kammy Algiers, 4.1.5: The Light-dependent Reactions, Biology Library, Libretexts project
(https://bio.libretexts.org/Bookshelves/Botany/Botany_(Ha_Morrow_and_Algiers)/04%3A_Plant_Physiology_and_Regulation/4.01%3A_Photosynthesis_and_Respiration/4.1.05%3A_The_Light-dependent_Reactions)

{kind=link}