
第四部 生命の進化
第7章 動物の発展【4億7500万 ~ 3億年前】
2.節足動物の時代【3億5000万 ~ 3億年前】
動物の大きな体は多くの場合、捕食に対する防御策であり、大多数の動物は体を可能な限り大きくしようとする。しかし、その大きさには当然ながら物理的な制限がある。そこで、動物の大きさを制限する要素は何かを考えてみる。陸生の節足動物(クモ、サソリ、ヤスデ、ムカデ、昆虫、その他の少数派グループ)の場合、その体制が持つ二つの特徴が大型哺乳類並みのサイズになることを制限していることが分かる。
- 一つ目の特徴は『外骨格』である。外骨格はほとんどがキチン質(注121)でできており、その強度の問題とスケール特性から、昆虫であろうとクモ形類であろうと、人間程度に巨大化しただけでも歩行すれば脚が折れ、体が崩れ落ちるだろう。
(注121)エビ,カニなどの甲殻類の殻やカブトムシのような節足動物の外骨格、軟体動物の器官部、カビ・酵母・キノコなどの真菌類の細胞壁などをつくっている物質の一般的呼称。化学的組成としてはキチンのほか蛋白質を含み、その比率は種類によって異なる。
- 節足動物が大きくなれないもう一つの理由は『呼吸器』である。昆虫やクモ形類は、体の最奥部までどれだけ酸素を行き渡らせられるかで大きさが制限されると考えられる。
現在、昆虫の体長は大きいものでも15センチ程度である。しかし、過去にはそれよりもはるかに大型の昆虫が存在した。それは、地球史上で最も酸素濃度の高かった時期であった。
石炭紀からペルム紀にかけての高酸素状態
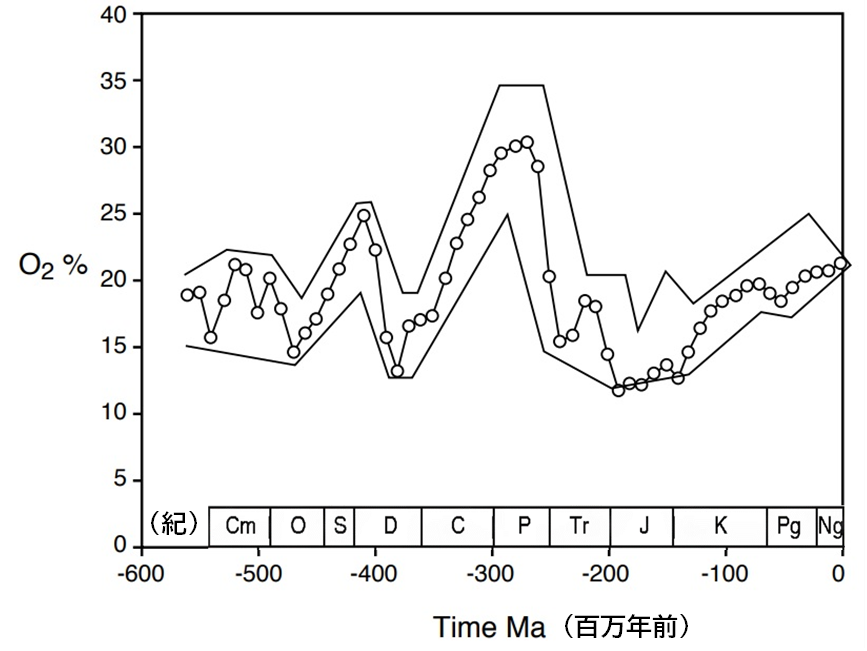
多様なモデルが示す過去の大気組成を検討すると、微細な数値の差異があるものの、 おおよそ3億2000万年前から2億6000万年前の期間における酸素濃度の極端な高さについては一致している。この時代の終わりには酸素濃度がその最大値を記録した(<物語り その10>図114を右に再掲)。これらの石炭紀とそれに続くペルム紀の初めの時期が高酸素の時代であり、その影響は当時の生物相にも明確に現れている。その中でも、昆虫がその証明となる最も明確な証拠となる。
石炭紀の高酸素については、ニック・レーンが2002年の著書『生と死の自然史-進化を統べる酸素』(西田睦監訳、遠藤圭子訳、東海大学出版会、2006年)で見事に説明している。『ボルソーバーのトンボ』と題された章には、1979年に発見された化石トンボの翼幅(翅を広げた長さ)がおよそ50センチだったとある。同じ石炭紀の化石からは、翼幅が76センチにもなるものも発見されている。これも同じくとんぼの仲間で、『メガネウラ(巨大な翅脈の意)』(注121)という適切な名前が付けられている。翅だけではなく、胴部もそれに比例して大きく、幅は約2.5センチ、長さは30センチに達した。これは大まかにカモメと同じ大きさである。カモメは決して「巨大」な生物とは形容されないが、翼幅が50センチに達する昆虫は間違いなく「巨大」と言える。現代のトンボの中には、翼幅が10センチに達するものもいるが、それよりも小さいものが一般的である。

(注121)「メガネウラ」は約3億年前の石炭紀の森に生息した大型のトンボ類の一属で、翅が30cm以上の長さをもつ巨大昆虫として知られている。学名[Meganeura]は古代ギリシア語[megas・メガース(大きな)]と[neuron・ネウロン(腱)]からなる合成語で、メガネ・ウラと分けて読むのは誤りである。neuronとはもともと「腱」を意味する言葉で、その後の転用により現代語「ニューロン」とも同源となっている。ここでは、昆虫の翅に見られる中空のすじで、気管や神経を通す「翅脈」を指しており、「巨大な翅脈を持つもの」という意味で命名された。
この時代のほかの巨大生物には、翼幅48センチのカゲロウ、脚の長さが46センチのクモ、全長180センチを超えるヤスデやサソリがいた。体長1メートルのサソリは、その重さが20キログラムを超えてもおかしくはなく、両生類を含むすべての陸上動物を捕食する可能性のある恐ろしい存在であっただろう。ただし、両生類も独自に巨大な種をいくつか進化させていた。
昆虫の場合、呼吸器系が酸素を体の内部までどれだけ効率よく運べるかによって、その大きさの上限が決まる。すべての昆虫は、気管と呼ばれる細い管から成る呼吸器系を持っている。空気は能動的に気管へと取り込まれ、その後各組織に拡散する。空気を気管内に引き込むためには、腹部を定期的に伸縮させたり、翅を羽ばたかせたりして、気管の入口近くで空気の流れを作り出す。どちらの方法も気管系の効率を向上させる役割を果たす。飛行昆虫は動物界で最も代謝率が高く、酸素濃度を高めるとトンボの代謝率がさらに上昇することが実験的に確認されている。こうした研究から、トンボの代謝率、そしておそらくはそのサイズも、現在の21パーセントという酸素濃度によって制限されていることが明らかになる。

酸素濃度が節足動物のサイズを左右するかどうかは、長らく議論が続いてきた。酸素が大きさを決定すると示す最も信頼できる根拠は、『端脚類』(注122)の研究から得られている。
(注122)節足動物甲殻綱軟甲亜綱に属する端脚類は、体が左右に扁平で、頭部、八胸節、七腹節から成る。眼に柄がなく、体長は0.5~2cmが一般的である。一部は淡水や陸上で生息するが、大部分は海洋で生息する。ハマトビムシやワレカラなど、約5700種が存在する。
端脚類は小型の水生節足動物で、現代の海洋や湖沼に広く分布している。ベルギーのゴティエ・シャペルとイギリスのロイド・ペックは、多様な生息地から2000点以上の標本を採取し、溶存酸素量が多い水域に生息する端脚類が大型であることを発見した。アリゾナ州立大学のロバート・ダドリーは、より直接的な実験を行い、酸素濃度を高めた環境でショウジョウバエを飼育した。その結果、酸素濃度が23パーセントの環境では、世代ごとに体が大きくなることが確認された。少なくとも昆虫にとっては、酸素濃度を上げれば比較的短期間で体が大型化することが確認された。
巨大トンボが生まれた背景には酸素だけでなく、大気圧も影響を与えていた。大気圧自体が現代よりも高かったため、より多くの気体分子が大気中に存在し、巨大トンボはより大きな揚力を得ることができたと考えられる。
確かに、大気中に現在より多量の酸素が存在していたことは否定できない。問題となるのは、その理由である。
酸素濃度を左右する主要な要因は、還元された炭素や含硫黄鉱物(例えば『黄鉄鉱』)の埋没率であると、我々は既に見てきた(注123)。有機物が大量に埋没すると、酸素濃度は上昇する。そうであるならば、地球史上で酸素濃度が最も高かった石炭紀においては、炭素や黄鉄鉱が短期間で大量に埋もれたはずである。地層の調査により、その仮定が実際に正確であったことが確認されている。その証拠として、『石炭鉱床』が形成されているのである。
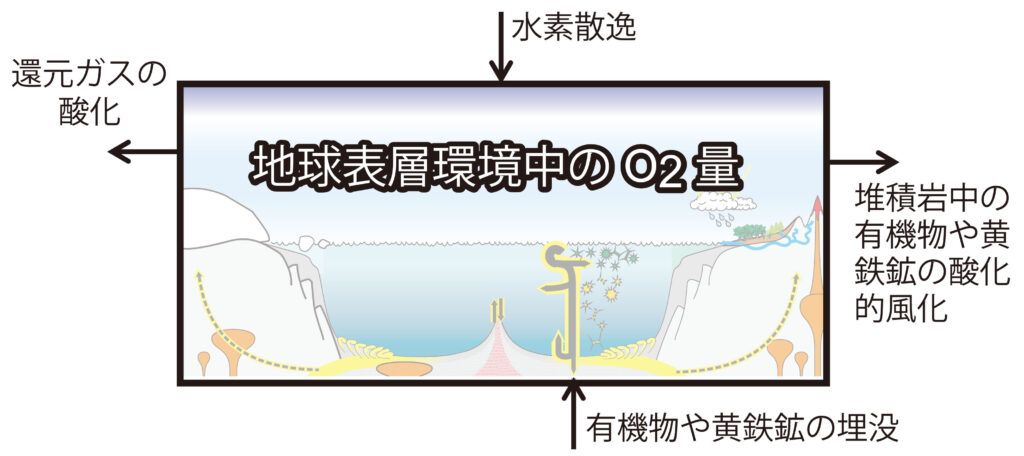
(注123)地質学的時間スケールでの地球表層環境中のO2量は、O2生成(「有機物や黄鉄鉱の埋没」と「水素の宇宙空間への散逸」(注124))とO2消費(「有機物や黄鉄鉱の酸化的風化」と「火成活動に伴って流入する還元ガスの酸化」)の収支で決まっている。ここでは光合成が直接的には含まれていないことに留意が必要である。それは、酸素発生型光合成により生成された有機物が完全に酸化分解されれば、正味でのO2生成はゼロになるためである。酸化分解を免れた有機物の埋没が正味のO2生成を担う。また、嫌気環境での有機物分解により発生する硫化水素が鉄と反応し、黄鉄鉱として埋没するプロセスも、正味のO2生成プロセスである。さらに、還元物質の酸化が地質学的時間スケールでのO2消費プロセスである。
(注124)地球における水素の大気散逸は、ジーンズ散逸[補注] (約10 – 40%)、電荷交換 (約60 – 90%)、極風 (約10 – 15%) によって発生しており、1秒あたり 3 kg の水素原子が散逸している。これに加えて、およそ 50 g 毎秒のヘリウムが、主に極風によって地球から散逸している。その他の大気成分の散逸は比較的少ない。
[補注] ジーンズ散逸 (Jeans Escape) は、静水圧平衡が成り立ち惑星に束縛されている大気からの散逸過程である。下層大気では密度が高く、高速で運動する気体分子はすぐに他の気体粒子に衝突・減速するために宇宙空間に脱出できないが、上層大気では密度が低いため他の気体分子と衝突せず、ある確率で脱出速度を超えて惑星重力から飛び去る。大気の「蒸発」とも例えられ、大気の「流出」と言われる流体力学的散逸とは区別される。
この期間は非常に長かった。3億3000万年前から2億6000万年前まで、つまり7000万年間にわたり、酸素濃度の高い状態が続いたのである(<物語り その10>図114参照)。地球の石炭鉱床の90%はこの時代の岩石から見つかっている。石炭紀には、石炭の埋没率が過去のどの時代よりも高かった。ニック・レーンの著書『生と死の自然史』によれば、これは驚くべき600倍もの高さであった。
しかし、ここで「石炭の埋没」という表現は不完全である。石炭とは実質的には太古の樹木の死骸であり、つまり、石炭紀には大量の倒木が急速に埋まり、それが後になって熱と圧力を受けて石炭となったのである。
石炭紀は、壮大なスケールで森林の埋没が起きた時代であった。さらに、石炭紀における有機物の埋没は陸上植物に限られたものではなかった。海中でも植物プランクトンや動物プランクトンの体内には炭素が大量に存在していた。プランクトンは海の森林のような存在で、その死後には、有機物に富む堆積物となって大量に海底に積もった。
こうした石炭紀特有の炭素蓄積が、他に類を見ない高酸素環境を生み出した。その根源的な原因は、偶然にも「地質学的なイベント」と「生物学的なイベント」が複数重なったことにある。
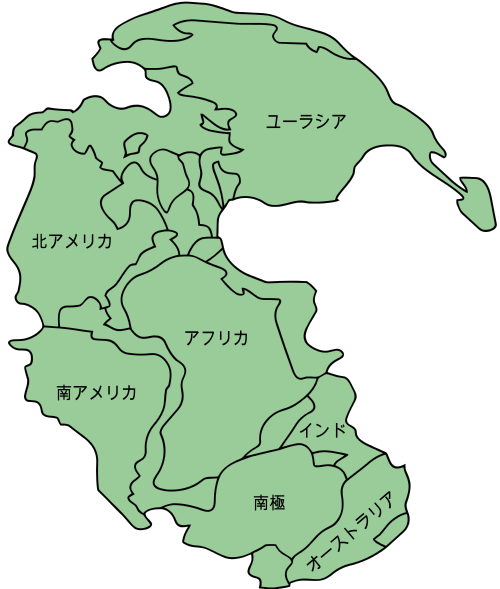
- まず、当時の大西洋が閉じて各大陸が合体し、一つの大きな大陸、『パンゲア超大陸』が形成された。ヨーロッパ大陸と北米大陸が衝突し、その後に南米大陸とアフリカ大陸も衝突した。その結果、大陸塊の接続部に沿って細長い巨大山脈が形成された。この山脈の両側には広大な氾濫原が出現し、山脈の位置による影響で広範囲にわたり湿潤な気候となった。大きな沼地周辺地域は次第に乾燥し、新たに誕生した樹木はその両方に進出した。
- 当時の樹木は、現代から見ると非常に風変わりだった。特に特筆すべきは、その根が非常に浅かったことである。そのため、大きく成長しても容易に倒れた(注125)。現代にも倒木は多いが、炭素蓄積という観点では石炭紀には遠く及ばない。湿潤な環境が植物の成長を促進したという以外にも、別の要因が存在していた。
(注125)石炭紀はデボン紀に多様化した植物が繁栄した時代であった。赤道地域の湿地にはシダ植物とシダ種子類が繁茂し、地球最初の森が形成された。リンボクやカラミテス(トクサ類)などの樹木のように大きくなるシダ植物が多く、やや乾燥した地域にはフウインボク(リンボク類)、メドゥロサ(シダ種子類)、プサロニウス(リュウビンタイ類)が分布していた。林床にはトクサ類やシダ類が繁茂していた。石炭紀とペルム紀の古生代後期は、これらのシダ植物とシダ種子類の時代と言える。リンボク類の茎(幹)を支えていた地下器官スティグマリアは丸太状で、この丸太状の器官からは根が出ていた。茎は葉を持つが、この丸太状の器官は葉を持たず根を生じる器官であり、「根を担う」という意味から『担根体』と呼ばれていた。担根体は種子植物には存在しない特徴であった。
- 約3億7500万年前に出現した森林は、本当に初めて「樹木」と呼べる存在から形成されていたのである。すなわち、これらの樹木は、リグニン(注126)やセルロースといった物質を使って体を支えていたのである。リグニンは非常に硬い物質ではあるが、現代では様々な細菌によって分解される。しかし、ほぼ4億年が経過した今でも、その分解には時間が必要である。単一の倒木が「腐る」までには何年もかかることがあり、また、軟木(スギやマツなど)よりリグニンの含有量が多い硬木では、より長い時間が必要となる場合も存在する。
(注126)リグニンは、高等植物の『木化』に関与する高分子のフェノール性化合物で、『木質素』とも呼ばれる。「木材」を意味するラテン語 lignum から名付けられた。成長するに伴い、高等植物の細胞壁に二次的に沈着し、一次壁は二次壁へと変化するが、この沈着物として最も顕著なものがリグニンである。リグニンは柔組織の細胞壁にも付加されるが、導管、仮導管、木部繊維において沈着が特に著しいため、その結果、細胞壁が肥厚する。この現象を「木化」または「木質化」と称する。この「木化」により、組織は強化され、安定性が向上する。
樹木の分解は、その炭素の大部分が酸化される過程で行われる。したがって、最終的に地面に埋もれたとしても、還元された炭素が地質記録に残ることは稀である。
- しかし、石炭紀には、樹木を分解する細菌がまだ十分に存在していなかったか、あるいは全く存在していなかった可能性がある。一般的な微生物は、植物の主要構成要素であるリグニンを分解することが難しいようである。そのため、石炭紀の樹木は倒れても分解されないままであった。時間とともに、この倒れた木はそのまま堆積物に覆われ、還元された炭素が埋蔵される過程で保存された。これらの樹木(や海洋プランクトン)が光合成を通じてどれだけの酸素を生成しても、この新たに生成された酸素が樹木の分解に使用されることはない。さらに、森林の木々は急速に成長し、倒れるので、酸素濃度は次第に上昇していった。
酸素と森林火災
石炭紀に酸素濃度がピークに達した影響は、動物の巨大化だけに留まらなかったに違いない。酸素は助燃性で、その量が多いほど火災は大きくなる。燃料の燃焼を促すからで、この場合の燃料は石炭紀の地球を覆っていた広大な森林であった。石炭紀には、過去最大級の『森林火災』(少なくとも6500万年前のチクシュルーブ小惑星衝突による森林火災が起きるまでは)が発生した可能性がある。少なくとも理論上は、酸素濃度が高いと火はより迅速に広がり、その勢いも強くなる。実際に、北米における石炭紀の堆積岩からは大量の木炭化石が発見されており、そのころには森林火災が発生していたことを証明している。その火災は現代よりも規模が大きく、強烈で、頻繁に発生していた。しかし、当時と現在では森林を構成する植物の種類が大きく異なるため、単純な比較はできない。
頻繁に発生する激しい森林火災に対抗するために、耐火性を持つ形態が進化していったと考えても不思議ではない。実際、植物は様々な耐火特性を発達させたことで知られている。例えば、樹皮を厚くする、維管束組織を深部に埋め込む(これはカンブリア紀に始まった)、ひげ根で茎を覆う鞘を形成するなどがそうである。
それほどの高酸素環境だったにもかかわらず、なぜ石炭紀の森林は一掃されるほど焼け落ちなかったのだろうか。この時代、火災は現代よりも頻繁に発生していたとはいえ、耐火性をもつ植物が登場し、植物自体が多くの水分を含んでいた上、無数の湿地が存在し環境全体の湿度が高かったため、火災の被害は抑制されたのである。
植物に対する高酸素の影響
動物と同様、植物も酸素がなければ生存できない。酸素は『光呼吸』(注127)の際に細胞内に取り込まれる。しかしほとんどの場合、その必要量は動物が必要とする量よりも大幅に少ない。
(注127)「光呼吸」とは、植物が光照射下において通常の呼吸(『酸化的リン酸化』(注128))と異なる方法で酸素 (O2) を消費し二酸化炭素 (CO2) を生成する現象である。状況によっては(CO2濃度が低い、高温などの条件下では)、光呼吸の速度が光合成の速度を上回ることがあり、すなわち、光呼吸による二酸化炭素の排出量が光合成による二酸化炭素の固定量を上回ることもある。
(注128)「酸化的リン酸化」とは、電子伝達系に結合して発生する一連のリン酸化(ATP合成)反応を指す。これは細胞内で起こる呼吸に関連した現象で、高エネルギー化合物のATPを生成する一つの経路である。好気性生物においては、エネルギー生成のための代謝の最終段階で、糖質、脂質、アミノ酸などの代謝がこの反応に集約される。ミトコンドリアの膜間腔とマトリックスの間で発生して水素イオンの濃度勾配のエネルギーを利用して、ATP合成酵素によってADPとリン酸からATPを合成する(54頁、図33を参照のこと)。
動物との違いをもう一つ挙げるとすれば、陸上植物の体の部位によって必要な酸素量に差が存在するという点である。多くの植物は二つの異なる環境で生存している。体の一部は空気中にあり、一部(根)は固体(土壌)中に存在している。根は地中に位置し、水分、固体、気体に囲まれ、地上部とは大きく異なる生息条件を有している。その結果、異なる進化を遂げる必要があった。葉は空気中に存在しているため、水分が失われることや日光が不足することを懸念する一方、過剰な水分により溺れることは心配しない。一方で、葉にとっては必要のない酸素量は根にとって概して必須である。低酸素状態では根系が真っ先に被害を受け、細胞が死滅することがある。根が存在する地中の環境は、大気中の酸素濃度が十分であっても低酸素状態に陥ることがある。特に、土壌に水分が多い場合、これはさらに顕著である。地下水の酸素濃度が低いと、根が窒息することがある。
逆に、酸素濃度が過剰な場合はどうかという問いについて、その関連データは限られているものの、既知の事実からすると、高酸素環境は植物に対して有害であると考えられる。大気中の酸素濃度が高い場合、光呼吸率が上昇する。さらに深刻な影響として、酸素濃度が高まると有毒な化学物質である『ヒドロキシル基』(注129)が増加する。これらの問題点をさらに検証するため、現在よりも酸素濃度を高めた密閉容器の中で様々な植物を育てる実験が行われた。
(注129)「水酸基」とも称される。-OHと表記され、化合物の名称には「ヒドロキシ」と表記される。アルコール性のもの、フェノール性のもののほか、遊離のもの(・OH)(ヒドロキシラジカル)が存在する。
実験結果によると、酸素濃度を35パーセント(石炭紀後期からペルム紀前期に見られた史上最高の酸素濃度とされる値)に上げると、純生産量(注130)は20パーセント低下した。したがって、石炭紀からペルム紀前期にかけての高酸素環境は、植物の寿命を一定程度短縮した可能性がある。しかし、化石記録からは、この期間に何らかの劇的な変化や大量絶滅が発生した形跡は確認できない。
(注130)植物の成長を測る指標の一つで、ある期間内の光合成による生産物(有機物)の総量を総生産量と称する。しかし、植物も呼吸を行うため、呼吸で消費される量を差し引いた残りが実際に植物体となり、これを「純生産量」という。
酸素と陸上動物
私たちの系統である脊索動物が陸上を征服するためには、陸地に適応するための大規模な改造が必要であった。
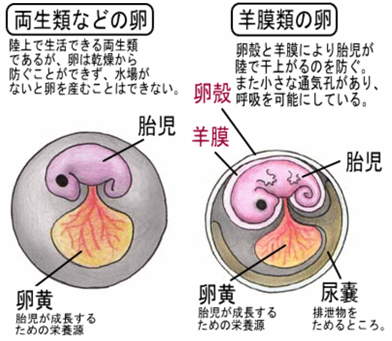
- 最も緊急の問題は、水中以外で卵の胚をどのように成長させるかであった。石炭紀後期からペルム紀の両生類は、おそらくまだ水中に卵を産んでいたため、湖や川のない陸地の資源を利用することはできなかった。
- この難題を解決したのが、『羊膜卵』と呼ばれるものの誕生である。今は爬虫類として知られる脊椎動物の系統が確実に存続できるようになったのは、この卵の存在によるところが大きい。
- 羊膜卵を有することにより、爬虫類、鳥類、哺乳類は祖先である両生類から区別されるようになった。
化石記録を見ると、羊膜卵を産む『羊膜類』(注131)は『単系統』であると推測される。つまり、同じ一つの祖先から派生し、この特徴が他の系統で独立に出現した形跡はない。その共通の祖先である両生類は、すでに石炭紀前期に存在していた。
- したがって、羊膜卵の出現という重要な出来事は、酸素濃度が上昇し始めていた時期に起こったと考えられる。最初の羊膜卵が形成された時、酸素濃度は現代と同程度か、それ以上に高かった可能性がある。
(注131)四肢動物のうち、発生の初期段階に胚が羊膜を持つものを指し、『有羊膜類』とも呼ぶ。両生類から石炭紀後期に分岐し、初期に『竜弓類』と『単弓類』の2系統に分化した。後に「竜弓類」の系統から爬虫類が、「単弓類」の系統から哺乳類が派生した。羊膜類が分岐し、爬虫類や哺乳類が誕生したのである。なお、羊膜類そのものは両生類には分類されない。
『爬虫類』は『単系統』と考えられており、約3億2000万年前の石炭紀前期に、祖先である両生類から分岐したと推定されている。その時期は、酸素濃度が上昇していただけでなく、両生類が陸生と水生に大きく分かれた時期でもあった。遺伝子の研究によると、この分岐が起きた時期は3億4000万年前まで遡ることが可能である。しかし、初めて爬虫類と認識される化石(陸生両生類ではない)は、世界の数箇所で、より新しい地層から発見されている。

ヒロノムスやパレオティリスといった小型爬虫類の化石は、石炭紀後期前半の化石化した木の根株に埋まっている状態で発見された。初期の爬虫類は非常に小さく、体長は10~15センチ程度のものが大部分であった。
初期の爬虫類には頭骨に鼓膜が存在しなかったため、聴力は弱かったか、もしくは全く聞こえなかった。また、迷歯類(注132)の両生類と異なり、大型肉食両生類によく見られるような大きな牙も保有していなかった。一方で、最初に真に爬虫類と称される動物は、頭部よりも後方の骨格に特徴が見られ、両生類よりも巧みに素早く動くことが可能であった。尾は体の大きさに対して長かった。
(注132)デボン紀末に初めて陸に上った四肢動物から起源し、石炭紀・ペルム紀・三畳紀にかけて繁栄し、その後衰退しつつも白亜紀前期まで存在した両生類の一派である。最も原始的なグループとして「イクチオステガ」(184頁、図135)が挙げられる。その名の通り、歯の表面のエナメル質が複雑に内部に折り込まれ、断面が迷路状になった「迷路歯」を特徴とする。これは「肉鰭綱」(シーラカンスなど)に属する祖先から受け継いだ形質である。
初の羊膜卵をこうした爬虫類が産んだのかどうかは、現在でも推測の域を出ない。ペルム紀前期の地層になるまで化石の卵は現われないし、その唯一の発見についても今なお見解が分かれている。
しかし、羊膜卵が誕生するまでには、両生類的な(乾燥を防ぐ膜がない)卵が陸上の湿った場所に産み落とされるという過程を経ていたと推定される。完全に陸上で産卵するためには、胚を包む膜(絨毛膜(注133)や羊膜)や、それを覆う皮革状の殻、あるいは多孔性で炭酸カルシウムの殻を生成する必要があった。
(注133)胎盤の胎児側の構成要素である。胎盤は胎児側の「胎盤胎児部」と母体側の「胎盤子宮部」から成り立っており、「絨毛膜」は胎盤胎児部の主要な部分を形成するものである。両者の間には胎盤腔(絨毛間腔)が存在する。
結果として、陸上で生存能力のある卵を首尾よく生み出す能力が発達した。
- この『羊膜卵の誕生』には、酸素濃度の増加や高温が一因であったと考えられる。
- 「卵生」という生殖戦略を採用する陸上動物にとって、生殖は大きなトレードオフをもたらすものである。卵の殻の孔は水分を保持するために可能な限り小さくし、数も減らすべきである。しかし、その結果として卵の中に酸素が入りにくくなる。
酸素がなければ卵は成長できない。羊膜卵の出現が高酸素時代と一致したのは偶然ではない可能性が高い。様々な環境で生活する動物が、今日も昔もこの生殖戦略を採用しているのも理解できる。この種の動物は大気中の酸素濃度の影響を受けており、酸素濃度が高いほど胚の成長は速まる。『胎生』も高酸素環境によって可能になったとする見解が存在し、低酸素環境では胎生は起こり得なかったと指摘する研究者もいる。その理由としては、少なくとも哺乳類では、胎盤を介して供給できる酸素量が母親の動脈血に含まれる量にも満たないためである。
四肢を持つ脊椎動物は魚類から進化したため、新たに直面した構造上の課題を多数克服しなければならなかった。
- 体を浮かせてくれる水がなくなり、陸上では四本の脚で体を支えながら移動しなければならないため、肩部や腰部の骨格の構造を全面的に見直し、移動を可能にする筋肉も一緒に発達させる必要があった。
- 同様に大きな問題となったのが、運動を持続できるだけの酸素をどのように確保するかであった。
- 初期の四肢動物は、運動と呼吸を同時に行うことはできないような体制で、同一の筋肉群を運動と呼吸の両方に利用していたとされる。
魚類の場合、運動を持続するのも、運動しながら呼吸するのも問題ないように見える。つまり、酸素の有無によって日常の活動が制約されることはない。
しかし、陸上の四肢動物は異なる。最初の形態は地を這うような姿勢で、脚は体幹部の側面から生えていた。このような体の構造で歩いたり走ったりすれば、体幹部が波打つように左右に曲がることになる。左脚を踏み出すと、胸部の右側と中の肺が圧迫され、右脚を踏み出すと左側に同じことが起きる。
このような胸部の歪みが発生する動きにより、通常の呼吸が可能とはならない。次の一歩を踏み出す前に、息をつく必要が生じる。しかし、これでは走行中の呼吸が困難である。したがって、現代の両生類や爬虫類は走行中に呼吸することができず、古生代の祖先も同様の問題を抱えていたことはほぼ間違いないと考えられる。速く走れない爬虫類が存在しないのはこの理由により、また、両生類と爬虫類が伏せ待ち状態で獲物を捕食するのも同じ理由による。走って獲物を追い詰めることはないのである。
現代の爬虫類の中でも、走行性能が最も優れているのはコモドオオトカゲであるが、獲物を追って全力疾走できるのはせいぜい9メートル程度である。
- この問題は、発見者である生理学者デイヴィッド・キャリアの名にちなんで『キャリアの制約』と呼ばれている。迅速な移動と呼吸を同時にできないというジレンマは、陸地にすみつく上で非常に大きな障害となった。
最初の陸生四肢動物は、サソリのような陸生節足動物に対してさえかなり不利な立場にあっただろう。その理由は、
- 脊椎動物の動くスピードは遅いうえに、呼吸のために頻繁に立ち止まる必要があったはずだからである。
- ここに酸素濃度が重要だったと主張する根拠が存在する。高酸素環境が存在したことにより、最初の陸生脊椎動物は陸上で繁栄する機会をつかむことができたのである。
呼吸問題から派生した影響の一つとして、初期の両生類や爬虫類が『三室心臓』(注134)を進化させたことが挙げられる。この種の心臓は現代の両生類や爬虫類の大部分に見られ、移動中に適切な呼吸が行えない生物にとって有用である。獲物を追っている間は呼吸を行わないトカゲは、肺への血液供給に必要な第四の心室を持つ必要がない。トカゲは三つの心室を用いて全身に血液を送っているが、その反面、活動終了後の血液再供給に時間がかかるという問題を抱えている。

(注134)心腔が三つある心臓のことで、哺乳類や鳥類は四室を有する。両生類は心室は1つなので、動脈血と静脈血が混じり合い、酸素と二酸化炭素の交換という点ではあまり効率がよくなく、肺呼吸のほかに皮膚呼吸も行っている。爬虫類も2心房1心室だが、心室を2つに分ける壁が不完全ながらも存在する。両生類よりも肺と心臓が強くなったため、皮膚呼吸をする必要がなくなり、より陸上生活に適応しているといえる。なお、鳥類と哺乳類は、左右の心房と左右の心室の4つの部屋に完全に分離された2心房2心室の心臓を持つようになり、動脈血と静脈血が混ざらないので、効率よく酸素と二酸化炭素の交換を行うことができる。
酸素と気温、生殖と体温調節
ここで、陸生動物が選択できる生殖方法を概説し、それを酸素濃度および気温の問題と関連付けて考える。先に見たように、
生殖には「卵生」と「胎生」の二つの戦略が存在する。卵生の場合、卵は炭酸カルシウムの殻や、より柔らかい革のような殻で覆われる。現在、鳥類の卵は全て炭酸カルシウムの殻を持ち、卵生の爬虫類の卵の大部分が革(または羊皮紙)状の殻を持つ。残念ながら、どちらがより酸素を通しやすいかについてはデータが不足している。
卵生か胎生かは、陸上動物にとって大きな意義を持つ。胎生で育つ胚は、気温変動、乾燥、または酸素欠乏といった危険から保護される。その代わり、母親は 体重が増え、胎児の分まで食物を摂取することが必要になるうえ、間違いなく捕食者に狙われやすくなる。卵生ではその心配がない反面、胚を取り巻く環境(卵の内側)が胎生ほど安全ではないというデメリットがある。卵が捕食されたり、外界が致命的な状況になったりすることで、胚の死亡率が高くなるのである。
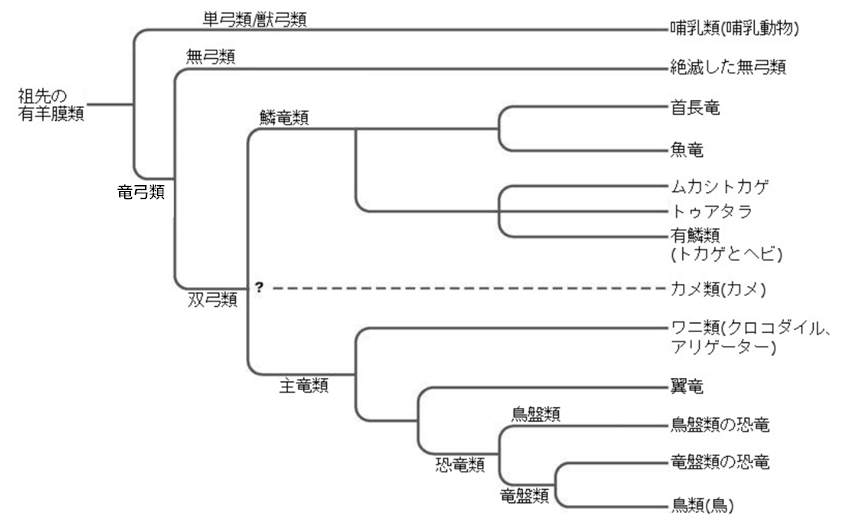
図144「有羊膜類の進化系統」に示されているように、石炭紀前期が終わる頃には爬虫類が『三つの大きな祖先系統』に枝分かれし、それぞれが独立した分類群になっていた。一つは『哺乳類』に(単弓類)、もう一つは『カメ類』(注135)に(竜弓類-双弓類)つながり、後の一つはまた別の『爬虫類』に(竜弓類-双弓類-鱗竜類/主竜類)なってのちに『鳥類』を生んだ。
(注135)頭蓋骨の眼窩後部両側に「側頭窓」と呼ばれる穴を持たないことが一つの特徴であり、かつてカメも「無弓類」に分類されていた。しかし、「側頭窓」を持たない生物全てが「無弓類」に分類されるわけではなく、DNAの解析結果からカメは、ワニや鳥類など主竜類に近縁であることが明らかになった。また、石灰質の卵殻を持つこと、発達した腹肋骨など他の無弓類には見られない特徴を持つなど、解剖学的な面からも無弓類とは別系統であるとされている。
化石からは、三つの主要な群体が様々な種から構成されていることが明らかになる。その多彩な化石記録から、これらの分類群がどのように進化してきたかが次第に明確になってきている。その結果、
『爬虫類』という概念そのものの再評価が迫られているのである。通常、爬虫綱には現生の「カメ類」、「トカゲ類」、「ワニ類」が含まれている。しかし、「爬虫類とは何か」という問いではなく「爬虫類とは何でないか」によって最近では定義されるようになっている。つまり、
- 「鳥類や哺乳類に固有の特徴を持たない『羊膜類』」と定義されるのである。
- 一般的に認識されていない事実として、これら三つの系統(上記『三つの大きな祖先系統』)が誕生した時代には、氷河が広がり、酸素濃度が非常に高かった。それゆえ、「低温だが酸素の多い世界」で生まれたことが、これらの動物たちの生物学的特性に多大な影響を与えたと考えることが可能である。
いくつか具体的に見てみると、
生命の歴史に関する長年の謎の一つに、動物の『体温調節』がどのように発達してきたかという問題がある。体温調節の方法は三つある。『変温性』、『内温性』、そして『恒温性』である。
- 「変温性」とは「外温性」とも言い、体温が主に外部環境によって制御される状態を指す。
- 「内温性」は体温が主に体内の代謝熱により維持される状態を指し、大型の動物に主に見られる。
- そして「恒温動物」は内温性の動物のうち、自律的に体温を制御している動物を指す。
これらがどのように進化したのかは長らく科学研究の焦点であった。特に注目され、多様な見解が飛び交っているのが「体温調節経路」についてであり、その中でも「恐竜が恒温動物であったかどうか」という問いが特に重要である。体温調節は生体内の生理機能によるところが大きいうえ、調節にかかわる体の部位(被毛など)がほとんど化石に残らないことが、議論を呼ぶ主な理由となっている。
現生の哺乳類や鳥類はすべて恒温性で、前者には体毛があり、後者には羽毛があることは広く知られている。同様に、現生の爬虫類はすべて変温性で、体毛も羽毛ももたないことも広く認識されている。絶滅した種については、まだ意見が一致していない。
ここで注目すべきは、酸素濃度や地球の気温が、過去の様々な祖先種の体温調節機能や特徴的な外被に影響を与えたかどうかである。
爬虫類の分化
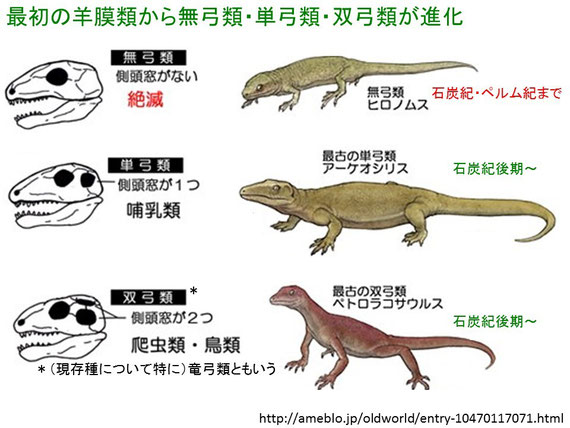
頭骨に開いた穴の数を数えると、『爬虫類』の三つの主要な祖先系統を容易に区別できる。『無弓類』の頭骨には側頭窓と呼ばれる大きな穴がない。『単弓類(哺乳類の祖先)』の頭骨には一つずつ穴があり、『双弓類(恐竜、ワニ類、トカゲ類、ヘビ類)』(注136)の頭骨には二つずつ穴がある。化石記録によると、これら三つの系統は酸素濃度が高い時期に出現したと考えられる。
(注136)頭蓋骨の両側に「側頭窓」と呼ばれる穴を、それぞれ2つ持つ。この穴ができたことでその下部の骨が細いアーチ状になっているが、これを解剖学では「弓」と呼ぶ。このため学名は「二つの弓 = 双弓類」と名付けられた。なお、片側に「弓」を一つ持っているものは「単弓類」、一つも持っていないものは「無弓類」と呼ばれる。
双弓類に属する最初の動物の化石は、石炭紀末の岩石から発見されている。その体長は20センチ程度であった(図147参照)。双弓類が誕生してから、酸素濃度が本格的に下がり始めるまでの間(ペルム紀中期から後期にかけての約2億6000万年前)、この群には大きな多様化や特殊化が見られなかった。石炭紀末からペルム紀前期にかけての酸素濃度が最高だった時期には、双弓類内部がいくつかのグループに分裂したにもかかわらず、双弓類自体は小型のまま、トカゲのような形状を保っていた。
- その後、双弓類から史上最大の陸上動物である『恐竜』が誕生し、中生代に地上を支配した。しかし、この段階ではそのような進化を予見する手がかりは一切なかった。
- 昆虫が高い酸素濃度の下で巨大化したのに対して、双弓類にはそのような現象が見られなかった。
- 関心を持つべきは、双弓類が恒温性だったかどうか、そして彼らがどのように繁殖を行ったかである。双弓類のどのグループについても、ペルム紀の決定的な証拠となる卵が見つからないため、その繁殖方法については確定的なことは言えない。ただし、皮膚状の殻を持つ原始的な羊膜卵を産んだと考えられている。とはいえ、胎生であった可能性も排除できない。ペルム紀末(酸素危機が深刻化し、最終的に史上最悪の大量絶滅に至った時期)になると、双弓類の多様化が加速し、その多種多様性が広く認識されるようになった。それは何と言っても、双弓類から恐竜が生まれたからである。

双弓類は動きやすい形態を進化させて駿足の肉食動物になったのに対し、もう一つの系統である無弓類は別の方向に向かった。無弓類は速度に拘束されることなく進化し、カメ類という形態を取った。それ以前には、「パレイアサウルス」という移動性の低い大型生物であり、ペルム紀後期の化石爬虫類の中では最大種であった。
第三の爬虫類系統は「単弓類」であり、これが私たち人間の祖先である。石炭紀後期からペルム紀にかけての酸素濃度が高かった時代において、双弓類はほとんど変化しなかったのに対し、単弓類は異なる進化を遂げた。

双弓類と同様に、最も原始的な単弓類の化石は石炭紀後期の岩石から発見されており、やはり当時の双弓類のようにこの哺乳類の祖先も小型で、おそらく形も生活様式もトカゲに似ていたと推測されている(図149参照)。初期の単弓類は、双弓類(およびその祖先である両生類)のように変温性であったと推測されている。その後、
単弓類は二つの主要な系統、
- ペルム紀前期のディメトロドンに代表される『盤竜類』と、
- そのあとに続く『獣弓類』(注137)
を生み出した。のちにこの獣弓類から哺乳類が生まれるので、獣弓類は『哺乳類型爬虫類』とも呼ばれる。双弓類とは異なり、単弓類は高酸素環境下で多様化し、酸素濃度が最 高に達した時期には、陸上で最大の脊椎動物になった。
(注137)「盤竜類」は、古生代石炭紀およびペルム紀に繁栄した陸生脊椎動物のグループで、四肢動物上綱・単弓綱の下位分類群の一つである。初期有羊膜類から進化した単弓類の初期グループで、ここから「獣弓類」が派生した。初期においては盤竜類と大差ないトカゲの様な姿であったが、その後期においては体毛や恒温性を獲得した。この恒温性の獲得は複数の系統(派生的なディキノドン類と獣歯類)で確認されている。「哺乳類」はその唯一の現生群である。

石炭紀末期には、盤竜類は外見も行動も現代の大型のオオトカゲやイグアナに類似していたと推測されており、四肢は体の側面から出ていたとされる。石炭紀末にはその一部の種はコモドオオトカゲに匹敵する大きさに進化し、強力な捕食者であった可能性がある。ペルム紀が始まる約3億年前には、すでに盤竜類が陸上の脊椎動物全体の七割あまりを占めていた。食性も多様化し、魚食動物、肉食動物、そして最初の大型植物食動物に分かれた。盤竜類は捕食する側もされる側も、全長が4.5メートルほどに達することがあった。『ディメトロドン』のように背中に大きな『帆』を有し、実際より体がはるかに大きく見えるものもいた。

盤竜類はまた、爬虫類が走りながら呼吸ができないという問題を部分的にまたは完全に解決した。これは体の姿勢を変化させた結果である。単弓類は、現代のトカゲのように四肢を横に突き出すのではなく、胴体の下に四肢を持ってくる方向に進化した。これにより、体が地面から高くなり、身をくねらせて肺を圧迫することがなくなった。少なくとも肺の圧迫は大幅に軽減されていた。四肢はまだ完全に胴体の下から生えてはいなかったが、最初の四肢動物に比べて明らかに改善されていた。ペルム紀中期に登場した獣弓類(図151参照)では、体をさらに高く保つ能力が進化していた。
帆は、石炭紀末期からペルム紀前期にかけて生息した肉食動物と植物食動物の両方に見られ、盤竜類の代謝機能を理解する上できわめて重要な手がかりとなっている。帆は、朝の早い時間に速やかに体温を上げるための仕掛けであったと考えられる。帆で朝日を受けることで、捕食者も被食者も大きな体を短時間で温め、迅速に行動することが可能であった。日光に頼らずに体内で熱を生成できれば、獲物を捕らえるにしても敵から逃げるにしても有利であるため、自然選択はその機能を獲得する方向に作用したと推測される。
しかし、酸素濃度が高かった時代には、哺乳類の祖先はまだ内温性を獲得していなかったことが明らかにされている。では、この特徴が現われた時期はいつであったのだろうか。この劇的な進化を達成したのは、盤竜類に続いて登場した『獣弓類』であったと考えられている。
ここでもう一つ指摘しておきたいのは、高酸素の時代は低温の時代でもあったということである。この時期には大氷河が存在していたことが明らかになっているうえ、両半球の極地においては、大陸も海も大部分が氷に覆われていたことが判明している。
獣弓類は主に肉食性のグループと植物食性のグループの二つに分けられる。
約2億6000万年前には南アフリカの氷が消えていたものの、その地域は超大陸パンゲアの中で比較的高緯度(南緯約60度)に位置していたため、気温は低かったと推測される。また、依然として高酸素時代の最中であり、酸素濃度が現在よりも高かったことは確かであるが、その状況は徐々に変わりつつあった。
ペルム紀が進むにつれ、酸素濃度が低下していった。やがて肉食動物と植物食動物の双方で大規模な二つの適応放散が起きた。およそ2億7000万年前~2億6000万年前の時代には、『ディノケファルス類』が陸上を支配していた。この時期には肉食動物として『アンテオサウルス』など、植物食動物として『モスコプス』などが存在していた。
ディノケファルスは非常に大型で、現代の陸生哺乳類並みのサイズに近く、最大級のものは象ほどの重さになったと考えられる。例えば、ディノケファルス類の中のモスコプス属は南アフリカで多数の化石が見つかっており、体高は5メートルに達し、巨大な頭部を持ち、前脚が後脚よりも長かった。これを狩っていたグループも同様の大きさであった。
その後、およそ2億6000万年前にディノケファルス類とその捕食者は大絶滅に見舞われた。
この絶滅については未だに解明が進んでいない。ディノケファルス類についても、その直後に現れて陸上を支配した最初期のディキノドン類やその捕食者についても、限られたデータしか存在していない。
『ディキノドン類』は植物食動物であり、2億6000万年前~2億5000万年前の時期に優勢を誇った。しかし、ペルム紀の大量絶滅により地球上からほぼ一掃された。ディキノドン類を捕食していたのは三種類の肉食動物である。ペルム紀末に死滅した「ゴルゴノプス類」、それよりやや多様な「テロケファルス類」、そして三畳紀になって最終的に哺乳類へと進化する「キノドン類」であった。

動物の大きさと酸素濃度
大気中の酸素濃度が30パーセントを超えて未曾有のレベルになった頃、昆虫は前例のない大きさへと進化した。石炭紀後期からペルム紀前期にかけての巨大トンボなどは、地球史上最大の昆虫であった。
この事実は単なる偶然である可能性も否定できないが、多くの専門家は「高酸素が昆虫の大型化を促した」と主張している。昆虫の呼吸器系は気管を通して体内に酸素を供給 するため、酸素濃度が高ければ昆虫の体が大型化しても十分な酸素が体内に供給されたと推測できる。
それでは、酸素濃度の上昇とともに大型化するという傾向は脊椎動物にも適用されるだろうか。新しいデータは、その傾向が確認できることを示している。
古生物学者のミシェル・ローランは2006年に、石炭紀からペルム紀にかけて(約3億2000万年前~約2億5000万年前)の様々な化石爬虫類の頭骨の長さと体長を測定した結果、どちらの値も酸素濃度の変動に連動して変化していたことを発見した。
- 石炭紀後期に酸素濃度が高まると、化石爬虫類のサイズも大きくなり、ペルム紀中期に酸素濃度が減り始めると小さくなっていったのである。
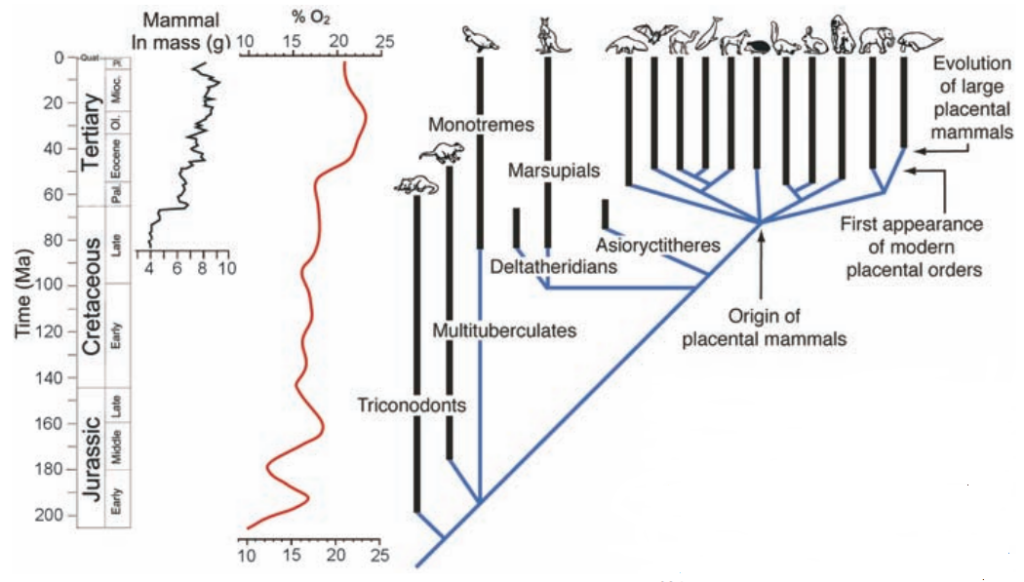
(注138)新生代の哺乳類に関する章でもう一度触れるが、アメリカの海洋学者ポール・フォーコウスキーらが後世の哺乳類について調べたところ、新生代前期にもこれとよく似た現象が起きていることがわかった。当時はやはり酸素濃度が著しく上昇したと見られていて、それと同時に哺乳類の平均的なサイズも増していたのである(注139)。
(注139)Paul G. Falkowski et.al., SCIENCE 309 (2005) 2202, “THE RISE OF OXYGEN OVER THE PAST 205 MILLION YEARS AND THE EVOLUTION OF LARGE PLACENTAL MAMMALS”

- 哺乳類型爬虫類における体の大きさが酸素濃度の上下とともに変動する傾向は、事実上確認されている。ペルム紀中期に生息していたディノケファルス類は、史上最大の獣弓類であり、酸素濃度が最高点に達した時期に出現したとされる。その後、酸素濃度が減少し始めると、ディノケファルス類に続く種々の獣弓類は頭骨が小さくなる方向に進化した。
- 特に注目すべきはディキノドン類である。ペルム紀末にはまだ比較的大型の動物(ディキノドン属や肉食のゴツゴノプス類)も存在していたが、すでにディキノドン類(図154参照)の多くは小型化していた。
ペルム紀末のシステケファルス属やディイクトドン属をはじめ、いくつかの属も体が小さくなっていた。
- 2007年の研究によれば、ペルム紀後期から三畳紀前期にかけて生息していたリストロサウルス属(注140)は、三畳紀のほうがペルム紀よりも小型であったとされている。これは酸素濃度が急減した時期と一致しており、同時期に生息していたキノドン類(図155参照)も体が小さかったという事実が同じ研究で明らかになっている。
(注140)約2億5,400万- 約2億4,800万年前(中生代三畳紀前期)のパンゲア大陸に生息していた植物食性の単弓類。獣弓目のディキノドン下目に属し、リストロサウルス科に分類される。
三畳紀のカンネメエリア属やトリティロドン属など一部の例外はあるが、全体として見ると三畳紀の獣弓類はペルム紀よりも著しく小さい。ワシントン大学のクリスチャン・シドーの最近の論文も、この急速な小型化を支持している。
つまり、ペルム紀末から三畳紀にかけては、陸上動物の体の大きさと酸素濃度の間に強い相関関係が認められる。この事実は、「四肢動物は高酸素の環境下で大型化し、酸素濃度の低下に伴って小型化した」という結論を導く。
「有羊膜類」の一つである「単弓類」から誕生した大きな二つの系統、「盤竜類」と「獣弓類」、その後者の「獣弓類」から哺乳類が生まれてきたので、獣弓類は『哺乳類型爬虫類』とも呼ばれる。この「哺乳類型爬虫類」に端を発して、酸素濃度の高低によりそのサイズの大小の影響を受けつつ、哺乳類時代が築かれて行くことになるが、それには、以下の三つの時代を経ることになる。
- 『第一の哺乳類時代』はペルム紀中にあり、この時期には獣弓類やその祖先である単弓類が繁栄した。厳密には、この時期の生物はまだ本物の哺乳類とは言えない。しかし、その進化の過程においては、彼らは哺乳類に近い存在であった。多種多様な生物が存在し、南アフリカでは一時期に50もの属が存在したという。通常、一つの属には数種から多数の種が含まれるため、種レベルで見ればその多様性はさらに高い。全体で150種以上あったと見積もられている。

現代の南アフリカの気候や緯度は、約2億5500万年前のゴンドワナ大陸南部に存在した時代と大差ない。現在の南アフリカには299種の動物が生息している。当時の大型植物食動物に代わり、ディキノドン類(図154参照)が草原で大量に生息していた。ライオン並みの大きさをもつゴルゴノプス類から、イタチほどのサイズの獣歯類まで、様々な肉食動物が生息していた。その食料となったのは草ではなく、低木のグロッソプテリスやシダ類であった。これが第一の哺乳類時代におけるアフリカの様子であった。
- 『第二の哺乳類時代』は三畳紀後期から白亜紀末までであった。しかし、この時期には恐竜が支配者であり、哺乳類は厳しい環境の中で生き延びることを余儀なくされていた。生態系の隙間で生存し、夜になると土中や木の間に身を隠す生活をしていた。体の大きさは大きくても家猫程度で、通常はそれより遥かに小さかった。
- 最後の『第三の哺乳類時代』は白亜紀から第三紀の境界の大量絶滅(<物語り その10>図113)の後であった。この時期には、今日私たちがよく知る科の動物が急速に増加した。ネズミのような小型の動物がチクシュループ小惑星の猛威を生き延び、初期の大型動物であるブロントテリウムやウインタテリウム(どちらもサイに似た動物)へと進化し、さらに私たちに親しい多くの哺乳類へと至ったのである。


図表
図139 メガネウラの化石
フリー百科事典『ウィキペディア(Wikipedia)』「メガネウラ」(https://ja.wikipedia.org/wiki/%E3%83%A1%E3%82%AC%E3%83%8D%E3%82%A6%E3%83%A9)
図140 短脚類の例
フリー百科事典『ウィキペディア(Wikipedia)』「短脚類」(https://ja.wikipedia.org/wiki/%E7%AB%AF%E8%84%9A%E9%A1%9E)
図141 地質学的時間スケールでの地球表層環境中のO2収支
東邦大学理学部、プレスリリース 発行No.919 平成30年10月5日
「“退屈な10億年”は飢えと酸欠の時代だった~ 地質記録と理論モデルの融合から得られた
太古の地球像 ~」
(https://www.toho-u.ac.jp/press/2018_index/20181005-919.html)
図142 パンゲア超大陸
フリー百科事典『ウィキペディア(Wikipedia)』「パンゲア大陸」(https://ja.wikipedia.org/wiki/%E3%83%91%E3%83%B3%E3%82%B2%E3%82%A2%E5%A4%A7%E9%99%B8)
図143 羊膜類と両生類などの卵構造の相違
古世界の住人
(https://ameblo.jp/oldworld/entry-10009215686.html)
図144 有羊膜類の進化系統
生物学 第2版 — 第29章 脊椎動物 —,OpenStax, Rice University,6100 Main St., Houston,
TX 77005-1827にて公開されている教科書“ Biology 2e”の日本語訳(https://medium.com/@BetterLateThanNever/%E7%94%9F%E7%89%A9%E5%AD%A6-%E7%AC%AC2%E7%89%88-%E7%AC%AC29%E7%AB%A0-%E8%84%8A%E6%A4%8E%E5%8B%95%E7%89%A9-bb7412be3b72)、図29.24に「竜弓類」を追記
図145 ヒロノムスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「ヒロノムス」(https://ja.wikipedia.org/wiki/%E3%83%92%E3%83%AD%E3%83%8E%E3%83%A0%E3%82%B9)
図146 両生類と爬虫類の三室心臓
ファーマスタイルWEB 2019年07月号
(https://www.credentials.jp/2019-07/medical-diagram-201907/)
Medical Diagram、106 心臓の進化の法則
図147 有羊膜類の頭蓋骨と動物例
古世界の住人
(https://ameblo.jp/oldworld/entry-10470117071.html)
図148 パレイアサウルス科スクトサウルスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「スクトサウルス」(https://ja.wikipedia.org/wiki/%E3%82%B9%E3%82%AF%E3%83%88%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9)
図149 アーケオシリスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「単弓類」(https://ja.wikipedia.org/wiki/%E5%8D%98%E5%BC%93%E9%A1%9E)
図150 ディメトロドンの想像図
絶滅動物図鑑「ディメトロドン」
(https://zetsumetsudoubutsu.com/dimetrodon.php)
図151 テトラケラトプスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「テトラケラトプス」(https://ja.wikipedia.org/wiki/%E3%83%86%E3%83%88%E3%83%A9%E3%82%B1%E3%83%A9%E3%83%88%E3%83%97%E3%82%B9)
図152 アンテオサウルスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「アンテオサウルス」(https://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%B3%E3%83%86%E3%82%AA%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9)
図153 モスコプスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「モスコプス」(https://ja.wikipedia.org/wiki/%E3%83%A2%E3%82%B9%E3%82%B3%E3%83%97%E3%82%B9)
図154 エオディキノドンの想像図
Wikipedia, the free encyclopedia“Eodicynodon”
(https://en.wikipedia.org/wiki/Eodicynodon)
図155 キノドンの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「キノドン類」(https://ja.wikipedia.org/wiki/%E3%82%AD%E3%83%8E%E3%83%89%E3%83%B3%E9%A1%9E#:~:text=%E3%82%AD%E3%83%8E%E3%83%89%E3%83%B3%E9%A1%9E%EF%BC%88%E3%82%AD%E3%83%8E%E3%83%89%E3%83%B3%E3%82%8B%E3%81%84%E3%80%81Cynodontia,%E5%8B%95%E7%89%A9%E3%81%AE%E4%B8%80%E7%BE%A4%E3%81%A7%E3%81%82%E3%82%8B%E3%80%82)
図156 酸素濃度の上昇と哺乳類の平均的なサイズの増加の相関
Paul G. Falkowski et.al., SCIENCE 309 (2005) 2202, “THE RISE OF OXYGEN OVER
THE PAST 205 MILLION YEARS AND THE EVOLUTION OF LARGE PLACENTAL MAMMALS”
図157 ディノケファルス類のエステメノスクスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「ディノケファルス類」(https://ja.wikipedia.org/wiki/%E3%83%87%E3%82%A3%E3%83%8E%E3%82%B1%E3%83%95%E3%82%A1%E3%83%AB%E3%82%B9%E9%A1%9E)
図158 顕生代の地質年代区分
『ウィキワンド(Wikiwand)』「地質時代」(https://www.wikiwand.com/ja/%E5%9C%B0%E8%B3%AA%E6%99%82%E4%BB%A3)
図159 グロッソプテリスの想像図
Wiki Prehistorica
(https://prehistoricearth.fandom.com/wiki/Wiki_Prehistorica)
(https://prehistoricearth.fandom.com/wiki/Glossopteris)
図160 ブロントテリウムの想像図
絶滅動物図鑑「ブロントテリウム」
(https://zetsumetsudoubutsu.com/brontotherium.php)
図161 ウインタテリウムの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「ウインタテリウム」(https://ja.wikipedia.org/wiki/%E3%82%A6%E3%82%A4%E3%83%B3%E3%82%BF%E3%83%86%E3%83%AA%E3%82%A6%E3%83%A0)
