
第13章 人類と10度目の絶滅【250万年前 ~ 現在】
数十年前、世界が新たな大量絶滅の時期に突入していると指摘する本が何冊か出版された。その中の一冊であるリチャード・リーキーの『六度目の絶滅(The Sixth Extinction)』では、これまでに説明してきた『ビッグファイブ』について明示している。ビッグファイブとは、全体の50パーセント以上の種が失われた大量絶滅を指し、具体的にはオルドビス紀、デボン紀、ペルム紀、三畳紀、白亜紀の末に起きた事象を指す(<物語り その10>図113を参照)。
しかし、私たちは実際には大規模な大量絶滅が10回起きたと考えている。これらは全て、前章で見たPETMやジュラ紀、白亜紀に起きた小規模な絶滅とは明確に区別されるべき事象である。10回の大量絶滅は年代順に次のようになる。
- 大酸化事変による絶滅(<物語り その7>「大酸化事変とスノーボールアース」節を参照)
古原生代のはじめ、約24億年前に起こったこの大規模な事象は、死滅した種と個体数の割合から考えて、最も壊滅的なものだったかもしれない。当時生息していたほとんどの微生物にとって、酸素は有毒であったはずだ。さらに、最初の『全球凍結(スノーボールアース)』が同じ時期に起きたことから、これは史上最初で最も悪影響を及ぼした絶滅であったと考えられる。その結果、微生物が呼吸することができる空気がなくなった。空気自体は存在していたが、その性質は完全に変わっていた。これは水生生物にとっても同じで、酸素という名の毒ガスが海を満たしていた。 - クライオジェニアン紀の絶滅(<物語り その8>「生命とスノーボールアース現象」節を参照)
原生代末に『スノーボールアース現象』が連続して二度起き、塵で汚れた黒い氷が海と陸を厚く覆った。光合成は大幅に減速し、ほぼ停止した。数多くの多様な生物(海中の生物が圧倒的に多かった)が死滅した。多様性だけでなく、生物全体の生物量(バイオマス)も大幅に減少した。 - エディアカラ紀後期の絶滅(<物語り その8>「世界に点在するエディアカラ生物群」節を参照)
ストロマトライトや微生物マットなど、原生代から古生代にかけて生息していたエディアカラ生物群がこの時期に絶滅した。『エディアカラの園』を荒らした生物は食欲旺盛で、さらに注目すべきは、これらの生物が活発に移動していたことである。それらは何でも食べ、動きの遅い微生物で覆われた海や陸を飽くことなく食べつくした。 - カンブリア紀後期のSPICEイベントによる絶滅(<物語り その9>「カンブリア紀の終焉 - 顕生代初の大量絶滅」節を参照)
『三葉虫』の大部分、バージェス頁岩の多様な『奇妙奇天烈動物』、その他多くの生物がこの期間に絶滅した。特に重要なのは、この時期を境に三葉虫の形態が大きく変化したことである。それまでの三葉虫は体節も目も原始的で、ほとんど防御の役に立たない装飾的な特徴にすぎなかったし、防御の姿勢をとるために体を丸めることすらできなかった。この変化の主な原因は、捕食者の増加だったと考えられる。頭足類の『オウムガイ類』は、真に大型と称することができる最初の肉食動物で、動きが速く、防御装備を備えていた。この化学的な変化と同時に、オウムガイ類の出現もこの絶滅に寄与したであろう。 - オルドビス紀の大量絶滅
オルドビス紀の大量絶滅とは、約4億4400万年前のオルドビス紀末に、三葉虫、腕足類、ウミリンゴ、サンゴ類、筆石、コノドントなど、それまで繁栄していた熱帯の生物が大半が絶滅した事件である。これにより、当時生息していた生物種の85%が絶滅したと推定されている。絶滅の原因としては、冷涼化または海水面の変動が考えられている。 - デボン紀の大量絶滅(<物語り その10>参照)
海底および海中に生息する動物が絶滅した。これを『最初の温室効果による絶滅』と呼ぶこともある。 - ペルム紀の大量絶滅(<物語り その13>参照)
温室効果が原因で陸海の生物が絶滅したとされている。 - 三畳紀の大量絶滅(<物語り その13>参照)
温室効果が原因で陸海の生物が絶滅したとされている。 - 白亜紀-古第三紀境界の絶滅(<物語り その15>参照)
温室効果と隕石衝突が絶滅の原因であった。 - 更新世末期~完新世にかけての大量絶滅
私たちと同じホモ属としては最古の種『ホモ・ハビリス』が登場した約250万年前から現在まで。気候変動と人間の活動が原因である。
我々が真剣に懸念すべきは、これらの絶滅の中で最新の第10回目の絶滅である。
10度目の絶滅へ
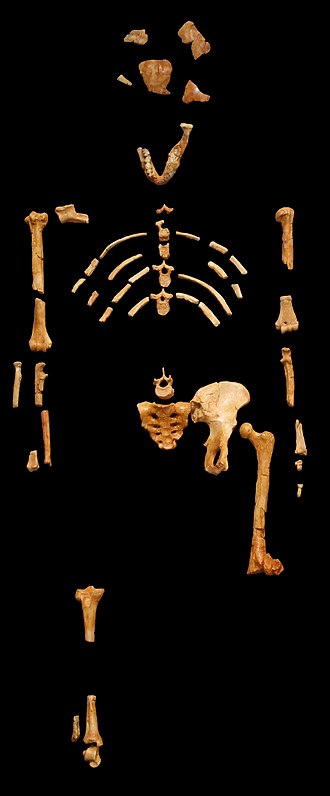
2010年、エチオピアから始まった巡回展で、非常に有名な化石がアメリカに運ばれた。これは最初のヒト科動物、通称『ルーシー』である。身長約107センチで、現存する骨は全体のわずか40パーセントに過ぎないが、彼女は我々に多くの情報を提供している。
一つの種の形態が雄と雌で異なることを『性的二形(雌雄二形)』と呼ぶ。これはヒト科に限られた現象ではないし、大きな体型が常に雄であるわけではない。例えば、多くの頭足類(オウムガイが例外)をはじめとする多くの動物では、雌が大型である。これは、卵を生産する器官が精子を生産する器官よりも大きな空間を必要とするためであると思われる。一方、ヒト科では一般的に雄が大きい。人間では、男女の体格には統計的に有意な差があり、人種によっては女性の身長は男性の約90~92パーセントとなっている。しかし、ルーシーの種であるアウストラロピテクス・アファレンシスの場合、これとは異なる状況が見られた。
ルーシーだけではなく、彼女と同じ種の他の個体の化石も発見されている。彼女が属する『アウストラロピテクス・アファレンシス(アファール猿人)』については、ドナルド・ジョハンソン率いる調査隊が1974年に彼女を発掘したときよりも格段に解明が進んでいる。最近では、同じ種の男性の骨格が発見され、しかも生前の身長を推定できるほどの良好な保存状態であった。この男性は『ビッグ・マン』と呼ばれ、身長は約152センチあった。
もしルーシーとビッグ・マンがアファール猿人の雌雄の典型であるなら、雌は雄の大体七割の大きさであったことになる。これほどの差は、行動や文化に何らかの影響を与えていたと思われる。
2012年には、ワシントン大学の人類学者パトリシア・クレイマーが二人の脚の長さから相対的な歩行速度を詳しく調べ、ビッグ・マンの最適歩行速度が時速4.7キロである一方で、ルーシーの場合は時速約3.7キロと比較的遅いことを明らかにした。この結果から、雌が雄に追従するのは非常に困難であったと推測され、捕食者の頻繁に存在する環境下では、常に息を切らしているのは生存にとって不利だったと思われる。そのため、クレイマーはヒト科の雌雄はチンパンジーのように一日の大半を分かれて過ごし、食物を探したり狩りをしたりする際には別々に行動していたのではないかと指摘している。
アフリカでは新たな化石の発見が続き、長年の学説が見直されつつある。ルーシーらの種がジオラマやイラストで再現されるとき、彼らは常に直立歩行をしているように描かれる。そしてその背後に広がるのは、小さな疎林と草原がモザイクのように絡み合った、アフリカ北部と東部の新鮮世後期の世界である。しかし、最近ではアファール猿人の雌の肩甲骨(ルーシーより10万年前のもの)が初めて発見され、その特徴から彼らが地上だけでなく木にも登れることが示唆された。私たちの遠い祖先が樹上生活をどの程度行っていたのかは、これまでにも頻繁に議論されてきた。これは、木に登るための適応構造を確認する手段がなかったためである。この新たな見方によれば、『アウストラロピテクス属』が木から降りてきた時期は、現在の定説よりも遅かった可能性があるということである。

『ヒト科』の生物が地球に出現したのは新しい時期であるが、我々が所属する『霊長類』自体の起源は白亜紀に遡る。霊長類の祖先である『プルガトリウス』が白亜紀-第三紀境界の大量絶滅を生き抜いたことは、我々にとって幸運であった。最も初期の霊長類にはキツネザルの系統に属するものも存在していた。より進化した霊長類、すなわち最初の『真猿類』(注191)は、4500万年前にはすでにアジアの化石記録に姿を表していた。その中で最も古いものは中国で発見され、今では『エオシミアス』と呼ばれている。

(注191)「哺乳綱霊長目真猿亜目」に属する動物の総称。「霊長目」は「原猿類」と「真猿類」の2亜目に分かれ、後者は霊長目の特性をより濃く持ち、前者よりもあらゆる点で高度な進化段階にあることを示している。オマキザル、オナガザル、ヒトニザル(ヒト)の3上科から成り、さらにマーモセット科、オマキザル科、オナガザル科、ショウジョウ科、ヒト科の5科に分かれている。

約3400万年前には、エオシミアスより確実に知能が高く、体格が大きく、恐らくはより攻撃的な猿がいくつか出現した。その一つが『カトピテクス』で、頭蓋の大きさは小型の猿と同程度で、相対的に平坦な顔を持っていた。カトピテクスは、突然変異によって人類と同じ歯並び(上下左右にそれぞれ二本の切歯、一本の犬歯、二本の小臼歯、三本の大臼歯)をもつようになった最初の霊長類である。
現在、私たち自身の進化の系統樹についてはかなり解明されており、『人類』が最初に現われたとされる場所と時期、つまりアフリカにおける『アウストラロピテクス』の誕生までが明らかになっている。すなわち、
- 私たちが属する科は『ヒト科』と呼ばれ、その歴史はおそらく600万年前~500万年前に始まったと見られる。
- この時期に、前述のルーシーら『アウストラロピテクス・アファレンシス(アファール猿人)』が出現した。
以降、ヒト科の生物は九種が確認されている。ただし、この数については現在も議論が続き、過去の化石に対する新たな解釈や新たな発見が公開されるたびに、その数は変動しているようである。しかし、
- 更新世以前の初期のヒト科生物で最も重要なのは、道具を使用する能力からその名がつけられた『ホモ・ハビリス(「器用なヒト」の意)』である。我々と同じホモ属としては最古の種で、約250万年前に出現した。
- 約150万年前にはホモ・ハビリスから『ホモ・エレクトス』が生まれ、
- そしてホモ・エレクトスから最終的に我々『ホモ・サピエンス』が派生したが、それは約20万年前にエレクトスの直系の子孫として生まれたか、あるいは『ホモ・ハイデルベルゲンシス』という中間の段階を経て進化したかのどちらかと考えられている。
- ホモ・サピエンスはさらにいくつもの変種へと細分化した。『ネアンデルタール人』を変種の一つとして扱う研究者もいれば、「ホモ・ネアンデルターレンシス」という別の種と解釈する者もいる。
ネアンデルタール人のDNAについては、その復元と解読を試みる新たな研究が数多く行われており、人類古生物学の中でも特に興味深い分野となっている。
- 最新の証拠によると、現生人類と我々の現在のDNAが出現する前に、人類とネアンデルタール人の系統はすでに分岐していたとされている。
- したがって、ネアンデルタール人が我々から生まれたのでもなく、その逆でもない。どちらも、すでに絶滅した共通の祖先から進化したので、その祖先はどちらの種とも異なる存在であった。
化石記録によると、我々の種(より原始的な『ホモ・サピエンス』と区別するために『現生人類』と呼ばれることもある)の中で、現時点で知られている最古の仲間は、19万5000年前に現在のエチオピアにあたる地域で生活していた。
しかし間もなく、この集団はアフリカ大陸の遥か南へと旅立ち、それから北へと進み、アフリカからユーラシア大陸に渡り、次第に世界中に広がった。その結果、この放浪者たちは自らを他の仲間から遮断し、自分たちが辿り着いたそれぞれの環境に適応していくことになった。
環境条件は場所によって大きく異なった。たとえば、北部地域はアフリカの草原に比べて日光が少なく、氷に覆われていた。そういった条件下で生き残るには、形態的にも生理的にもこれまでとは全く異なる適応が必要であった。これは、その中間のすべての地域にも当てはまる。
我々の個体数が増加するにつれて種内の多様性も増大し、さまざまな進化的な変化が生じた。だがそれらすべては同一種内で発生したものである。
最後の氷河期と生命
長らく、気候学者たちは250万年前からの『気候変動』(氷床形成による海面低下とそれに伴う長い寒冷期と、それより短い温暖期が交互に現れる現象)が地球の軌道の変化による結果であると認識してきた。この観点を初めて提唱したのはミルティン・ミランコビッチであった。そして、これらの変化は徐々に進行するものだと広く認識されていた。しかしながら、氷床コアの入手可能性とそれにより得られる前例のない精度の気候データが新たな解釈を生む契機となった。
氷床コアの記録や深海生物、同位体に関するデータの参照により、過去80万年間に存在した『間氷期』(寒冷な氷期と温暖期の間の時期)の平均期間が約1万1000年であることが明らかとなった。これは地球の歳差運動(地球の自転に伴う微細な首振り運動により、自転軸の向きが約26,000年周期で変わる現象)の周期のおおよそ半分に相当する。現在の間氷期は既に1万1000年以上続いており、それが1万4000年に達している記録も存在する。したがって、この瞬間でも氷河が進行していると考えるべきかという疑問が生じるが、答えは明確に否定的である。それは、気候に影響を与える地球の運動要素が歳差運動だけに限定されないからである。記録によると、約45万年前から35万年前までの期間には、1万1000年を大幅に超える長さの間氷期が存在した。この間氷期は、地球の公転軌道の「軌道離心率」(注192)が最小であった時期と一致していた。そして、この離心率が最小化する現象は現在も進行中である。つまり、現在の間氷期はさらに数千年から数万年続く可能性があることを示している。
(注192)軌道離心率(e)は、軌道形状がどれほど円から逸脱しているかを示す値である。標準的な条件下では、軌道離心率の値により、円、楕円、放物線、双曲線が定義される。円の場合はe=0、楕円の場合は0<e<1、放物線の場合はe=1、そして双曲線の場合はe>1となる。
『更新世』では、約250万年前に重大な気候変動が始まった。氷河期が訪れる以前の時代、寒冷な気候の高緯度地域では広大な草原やツンドラが地表を覆っていたが、それが氷に取って代わられた。年々、雪と氷は増大し、氷河形成を引き起こし、その氷河は徐々に南へと進展した。結果として、大陸氷河と山岳氷河が融合し始め、その結果として、大地は氷と冬の支配下に置かれた。
しかし、地球全体が氷に覆われるわけではなく、熱帯地域やサンゴ礁は残され、一年を通じて日照が良好な温暖で快適な気候も存在した。しかしながら、おそらく、全く影響を受けなかった場所は地球上には存在しなかったと推察される。気候変動は全球的な範囲で生じ、風や雨のパターンに変化をもたらした。氷床から離れた場所でも気候変動の影響を受け、以前と比較して寒冷化したり、暖かくなったりする可能性があった。また、概して気候は乾燥する傾向にあった。
前進する氷河の前面では、寒冷で広大な砂漠地帯や半砂漠地帯が形成され、本来乾燥していたサハラ砂漠のような地域では逆に降水量が増えた。一方で、アマゾン川流域や赤道直下のアフリカに広がる広大な熱帯雨林は、氷河期が始まる前は数千万年間もの間気候が比較的安定していたにもかかわらず、明らかに気温が下がり乾燥化した。その結果、広大なジャングルが後退し、乾燥したサバンナに囲まれた島のようになった。
人類の拡散
気候変動の急速な進行の多くは、人類が世界各地に定住し始めた時期に発生したと見られる。おおよそ3万5000年前、進化の最後の微調整が施され、我々は現在の姿へと変化したと推定される。この新たな人類は『現生人類』と呼ばれ、世界の各地を少しずつ探索していった。彼らはゆっくりとではあるものの、堅実に新たな地域へと進出した。
この現生人類の移動は、徐々に地球全体に広がりつつある葉が一枚一枚落ちていくような過程であった。島大陸であるオーストラリアさえも、3万5000年前にはすでにホモ・サピエンスの居住地となっていた。それでもなお、北アジアは未発見のままであった。さらにアジアの彼方には広大な南北アメリカが広がっており、未だに人の足が踏み入れていなかった。
現在のシベリアという広大な土地に最初に到達したのは、旧石器時代の大物狩猟者であった。彼らは3万年前にこの地に足を踏み入れ、厳しい気候の中で生きるための伝統をすでに持っていた。シベリア東部の石器は、当時のヨーロッパ人の石器とはいくらか異なり、明らかに東南アジアの剥片石器の文化に影響を受けていた。それでも、彼らは大型動物を狩るための大きな槍先を製作する重要な技術を編み出していた。
人類が初めてシベリアに足を踏み入れたのは、気候が比較的温暖であった時期であった。冷涼期が終わり温暖期が始まったことが、本来は住むに適さない地域へ人類の拡散を促した可能性がある。しかし、人類がシベリアに到達した直後に地球は再び冷却を始め、2万5000年前には既に氷河期特有の現象が進行していた。
西ヨーロッパと北アメリカでは、大量の大陸氷河が南方向に無情に広がり、厚さ1500メートルを超える氷が広大な大地を覆っていた。しかし、シベリアは湿度が低かったために氷が形成されなかった。木々がほとんど生えていない凍てついた大地を、人類は東へと進んでいった。木材が極めて少なかったため、狩猟した動物の皮や骨が重要な資源となり、最大級の獲物である『マストドン』(注193)や『マンモス』の骨が住居に利用された。このような状況から、彼らは大型獲物を狙い、主にマンモスやマストドンを狩猟していたと推測される。
(注193)ゾウ目マムート科マムート属に属する、大型の哺乳類の総称。原始的なゾウ類でありマンモスや現生のゾウに似ている。約4000万年前から11000年前まで生存していた。

約3万年前から1万2000年前の間に、小さな波が次々と打ち寄せるようにして人類はアジアを横断し、『ベーリング地峡』に到達したと考えられる。その時期は長期の寒冷期が続いており、北米を覆う大陸氷床の面積が最大に達していた。氷が増加することにより海面が下がり、かつて水面下にあった広大な土地が露出した。これは移動を助ける経路となり、それまで分断されていた島や陸地をつなぐ役割を果たしたと考えられる。しかし、その後すぐに急速な温暖化が始まり、氷が融けるとともに海面も上昇した。このようにして気温は徐々に上昇し始めたが、カナダの大部分と現在のアメリカ合衆国の多くを覆っていた大陸氷床は、1万4000年前になってもまだ溶けている最中であった。
その後まもなく、新たな現象が発生し、氷の融解が加速した。大量の氷が消失し、氷河が海にまで広がらなくなると、現在のカナダとアメリカ北部の東西の海岸から氷山が分離しなくなった。
氷河期がピークに達していた時期(約1万8000年前~1万4000年前)には、毎春大量の氷塊が海に運ばれ、海水を低温状態に保つとともに冷たい風を生み出し、陸地を冷やしていた。しかし、氷山の生成が止まった結果、海風の温度が上昇し、大陸の氷は至る所で本格的に溶け始めた。
氷河が後退している最前線は異様な環境となり、その厳しさは尋常ではなかった。後退する氷河壁の周囲では絶え間なく強風が吹き荒れ、その風により運ばれた砂やシルトが大量に堆積し、レス(黄土)という地層を形成した。風によって運ばれてきた種子は、氷河前線に堆積した土壌で次々と生育し、最初はシダ類が生え、その後より複雑な植物が生えるようになった。ヤナギ、ビャクシン、ポプラなどの低木が初期の植生を形成し、氷河が支配していた環境に変化をもたらした。その後すぐに、場所によってさまざまな種類の植物が出現した。より温暖な西部では、トウヒなどの常緑針葉樹による低木林が成立し、より寒冷な大陸中部は永久凍土とツンドラに覆われた。しかし、どの地域においても、氷河が消えつつあるという事実に変わりはなかった。
氷河壁が融けながら北に移動する過程で、そのあとを追うようにツンドラが広がり、ツンドラのすぐあとには広大なトウヒの森が続いた。これらトウヒの群生は、北米の広範囲を覆い、疎林と密林の特徴を併せ持つ構造を有していた。密集した木々の間には、草地や低木が散在していた。これは、現在でも北西部に僅かに残る原生林に見られるような、ベイマツの大木が密生し、下草や朽ちた倒木が存在し大型動物や人間にとって非常に通り難い場所とは異なっていた。
氷河期を通じて、北米大陸の氷河の南側には様々な生息環境が存在した。これには森林ツンドラ、草原、砂漠が含まれ、大群をなす巨大哺乳類を養うための植物も生息していた。寒冷で氷の時代が世界の大半で終焉を迎えるにつれ、人間の数は顕著に増加し始めた。
一万年前には、人類は南極を除く各大陸に成功裏に定住し、そしてそれぞれの地域の環境に適応した結果、現在見られる多様な人種が形成された。
肌の色などの明らかな特徴は、日照量の違いに応じて生まれたにすぎないと長らく考えられていた。ところが最近の研究によると、人種に固有とみなされている特性の多くはそれぞれの環境への適応を高めるためというより、単に「性選択」(注194)によってもたらされたという可能性が浮上している。しかし、それ以外にも人種特有の特徴は発達していき、そのほとんどは形としては目に見えないものだった。
(注194)「雌雄淘汰」、「性淘汰」とも言われ、特定の性(通常は雄)の特異な形質を強化する選択圧のことを指す。クジャクの羽やライオンのたてがみ、シカの角や鳥のさえずりなど、雄に特有の形質の進化を説明するためにC.ダーウィンが提唱した。
アフリカは大型哺乳類の宝庫として称賛されている。これほど多様な大型の植物食動物や肉食動物は他の地域では見ることができない。しかしこの動物の楽園は特別な例ではなく、かつては一般的な風景であった。世界中の温帯や熱帯の草原地帯は、近年までアフリカの風景に酷似していた。しかし、様々な影響がカルー地方のゾウを絶滅に追い込んだように、ある異常な出来事により大型哺乳類の生物多様性は5万年前から急激に減少し始めた。
大型動物の絶滅は、絶滅を研究する科学者にとって大きな課題を提示する。しかし、過去から得た重要な教訓を考慮すると、大型動物の絶滅は小型動物の場合よりも生態系の構造に深刻な影響を及ぼすことが明確である。白亜紀末の絶滅が重要な意義を持つのは、多くの小型哺乳類が消滅したからではなく、非常に大きな恐竜が絶滅したためである。陸生の超大型恐竜が一掃された結果、陸上の環境は大きく変化した。同様に、過去5万年の間に世界各地で大型哺乳類の大部分が絶滅したことは、その影響が徐々に明らかになりつつある。そして、その余波が今後数百万年にわたって続くことが避けられないと見られている。
特に、更新世後期の約1万5000年前から1万2000年前の間に、北米に生息する大型哺乳類の大部分が絶滅し、少なくとも35の属(したがって最低でも35種)が消失した。このうち6種は、アメリカ以外では生存を続けたものの(南北アメリカでは絶滅したが、旧世界では生き残ったウマなど)、圧倒的多数が絶滅した。実際、絶滅は様々な分類群に及び、21の科と7の目が影響を受けた。これだけ多岐にわたる中で唯一の共通点は、絶滅したほとんど(もちろんすべてではない)が大型動物であったということである。
まとめると、更新世の絶滅は以下のような特徴を持つといえる。
- 絶滅したのは主に大型の陸上動物であった。比較的小型の陸上動物と、ほぼ全ての海洋動物は絶滅を免れていた。
- 大型哺乳類が最も多く生き残ったのはアフリカであった。北米では大型哺乳類の73%が失われ、南米では79%、オーストラリアでは86%だったにもかかわらず、アフリカでは過去10万年間で絶滅したのは14%にすぎない。
- 絶滅は主要な陸地で急激に起こったが、その時期は大陸により異なっていた。高精度の放射性炭素年代測定技術による調査から、数種の大型哺乳類については300年程度の間に完全に絶滅した可能性が指摘されている。
- この絶滅が新たな動物群(ヒト以外)の侵入によりもたらされたわけではないことは明白である。従来の学説では、高度に適応性を持った新種の生物が進化の過程で突然新しい環境に現れたとき、絶滅が生じる可能性が高いとされていた。しかし、氷河時代における絶滅については、この説明は当てはまらない。新たな動物相の到来が既存の種の絶滅と直接関連づけられる事例が一つも存在しないためである。
こうした様々な方面からの証拠を見て、大型哺乳類の大量絶滅の引き金を引いたのは人類だったと考える研究者は多い。一方で、氷河期末に生じた急激な気候変動が食糧源としての植物群を変え、それが絶滅の一因となったと主張する学者もいる。このように、
絶滅についての議論は主にその原因に焦点を当てており、大きな意見の分かれる二つの学派が形成されている。
- 過剰殺戮(人間による狩り)が主要因であると説く陣営
- 気候変動が主要因であると主張する陣営
である。
何が原因であれ、陸上生態系の大規模な再編がアフリカを除く全大陸で起きたことは間違いない。現在、そのアフリカも大型哺乳類を失いつつある。自然保護区だけが狩猟の対象となる動物の生息地となるにつれて、生息地が限られることで逆に密猟の標的となり易くなっている。
大型動物相の終焉がいつくるのかは定かではない。だがそれは、現在から未来を見ようとするからそう感じるのであって、実はすぐそこに迫っている。数千万年後、数億年後から過去を振り返れば、過去の1万年などは微々たるものであり、その間に何が起こったかを高度な技術でさえ正確には割り出せないだろう。今の時点では、大型哺乳類の時代が終わるのはまだ先のように感じるかもしれない。しかし、一旦それが過去の事実となってしまえば、時間の経過とともに一瞬で起こったように思えることが、時間の持つ奇妙な特性である。
現在、地球に生息する大型哺乳類は絶滅危惧種の大半を占め、それよりも小型の哺乳類の絶滅危機に瀕している種の数はさらに増加している。第四紀(約180万年前から現在に至る地質時代最後の期間)の大量絶滅の第一段階が大型哺乳類の消失だったとすれば、現時点では植物、鳥類、昆虫が次々と失われる段階にあると考えられる。原因は、地球上の太古の森林が農地や都市に変わってしまったためである。
図表
図240 化石人骨・ルーシー
フリー百科事典『ウィキペディア(Wikipedia)』「ルーシー (アウストラロピテクス)」(https://ja.wikipedia.org/wiki/%E3%83%AB%E3%83%BC%E3%82%B7%E3%83%BC_(%E3%82%A2%E3%82%A6%E3%82%B9%E3%83%88%E3%83%A9%E3%83%AD%E3%83%94%E3%83%86%E3%82%AF%E3%82%B9)
図241 プルガトリウスの想像図
古世界の住人「プルガトリウス」
(https://paleontology.sakura.ne.jp/purugatoriusu.html)
図242 エオシミアスの想像図
小田 隆 Oda Takashi@studiocorvo「エオシミアス 化石種」
(https://twitter.com/studiocorvo/status/840441482821681152?lang=zh-Hant)
図243 カトピテクスの想像図
古世界の住人「カトピテクス」
(http://paleontology.sakura.ne.jp/katopitekusu.html)
図244 ケナガマンモス(左)とアメリカマストドン(右)の比較
フリー百科事典『ウィキペディア(Wikipedia)』「ケナガマンモス(左)とアメリカ
マストドン(右)の比較」
(https://ja.wikipedia.org/wiki/%E3%83%9E%E3%82%B9%E3%83%88%E3%83%89%E3%83%B3)
