
第11章 ようやく訪れた第三の哺乳類時代【6500 ~ 5000万年前】
知られている中で最初の哺乳類、モルガヌコドンは、トガリネズミほどの大きさで、極めて痩せていた(注179)。2億1000万年前の三畳紀末期に存在し、大型の捕食者に怯えながら生活していたと思われる。このモルガヌコドンは、恐怖に見舞われつつも三畳紀-ジュラ紀境界(T-J境界)の大量絶滅を生き抜き、その後まもなく、原始的ながら真の哺乳動物がほかにも仲間に加わった。

(注179)モルガヌコドンは、哺乳類が祖先グループである単弓類キノドンから分岐した時期の生物であり、その存在期間は約2億年前の三畳紀後期である。モルガヌコドンは厳密には哺乳類ではなく、哺乳類を含むより大きなグループである『哺乳形類(ほにゅうけいるい)』に分類される。哺乳類の祖先グループの特徴と哺乳類の特徴を併せ持つ動物であり、哺乳類の系統図における基盤的な位置を占めている。この動物は、単純な歯しか持たない爬虫類とは異なり、奥歯に3つの出っ張りを持っていた。この出っ張りが噛み合わさることで、昆虫を効率的に噛み潰せた。このため、硬い甲殻を持つ昆虫も摂食できた。その後に現れる哺乳形類と哺乳類は、このモルガヌコドンに比べて大きな脳を持つと言われている。
ヒトを含む現生哺乳類はすべて、T-J境界絶滅をくぐり抜けたこの唯一の系統に由来する。
長かった恐竜時代が激烈な最後を迎えたのち、地球上では一変して、いわば「ネズミの大発生」が起こった。
哺乳類の進化は南から北へと広がっていったとされる。その証拠として南で新たに収集された化石が挙げられる。これらの化石は高度に進化した哺乳類のもので、北で発見されたどの化石よりも古い時代に属している。遺伝学の成果も同様の傾向を示している。DNAの比較や進化発生学(エボデボ)から得られた新たな知見が、この哺乳類の進化についても確認されている。特に注目すべき事実は以下の3つである。
- 第一に、哺乳類の主要な『分類群』(現生の18目および今も見られる亜目や科もいくつか含めて)は、実は恐竜の絶滅よりずいぶん前にすでに多様化していた。
これは長年の認識、すなわち哺乳類の分類群が白亜紀-第三紀(K-T)境界絶滅後に初めて進化したという認識を覆すものである。化石の記録からは、現代の分類群の多くがおよそ6000万年前、恐竜が絶滅した後に初めて出現したように見える。しかし、分子のデータは、これらの分類群が実際には約1億年前から多様化し始めていたことを示している。
- 第二に、初期の哺乳類の進化とそれに続く分岐は、前述の通り南の大陸で起こった。
- 第三に、遠縁と考えられていたいくつかの分類群が、実は近縁であることが明らかになった。
たとえばコウモリは、ツパイやヒヨケザルやサルと同じ上目(真主齧上目・しんしゅげつじょうもく)(注180)に属すると古生物学者たちは考えていた。しかし遺伝学的データにより、コウモリはブタやウシ、ネコ、ウマ、クジラの系統(ローラシア獣上目)に分類されることがわかった。同様に、クジラはアザラシ(鰭脚類・ききゃく類)につながる祖先系統から進化したのではなく、ブタに似た祖先(偶蹄類)から進化したことが明らかになっている。
(注180)「目」は生物分類のリンネ式階級分類における基本的な階級であり、その階級に属する分類群を指す。具体的には、「綱」の下、「科」の上に位置する。さらに、「目」の上に「上目」を設ける場合もあり、「目」の下には「亜目」、その下に「下目」、さらにその下に「小目」を置くこともある。
哺乳類の繁栄は解剖学的構造の変化によるところが大きい。
- 特に注目すべき変化の一つとして、「顎の骨と耳の骨の分離」が挙げられる。この変化により、哺乳類の頭骨は横方向と後方に広がり、脳が大きくなるという重要な進化を促したのである。
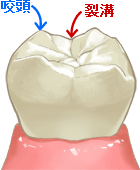
- しかし、これらの進化の中で最も重要とされるのが『歯の進化』である。モルガヌコドンの顎骨にある臼歯は上下が噛み合い、食物を細かく砕くことが可能であったという(注181)。
(注181)モルガヌコドンの歯列は大小三つの咬頭(臼歯の歯冠の尖っている部分)が漢字の「山」の様に直線で並んだ形態の臼歯が特徴的である。その上下が噛み合うことにより、ゴミムシのような硬い甲虫類を細かく砕き食べることができた。
現生の哺乳類は大きく二つに分けることができる。
- 一つは、祖先にあたる『有袋類』で、極小の子供を産み出し育児嚢で育てる特性を持つ。
- もう一つはその子孫で、種の数も個体数も有袋類より多い『有胎盤類』である。
近年のDNA研究では、有胎盤類は1億7500万年前には早くも有袋類から分岐し始めていたとの見解が示されている。この見解には化石による裏付けもあり、中国北東部の遼寧省で産出した化石は有力な証拠となっている。1億2500万年前の地層から見つかったこの完全な形の有胎盤類の祖先の化石は、DNA研究の結論を裏付けており、『エオマイア』と名づけられている。この化石の発見により、有胎盤類の最初の祖先がそれよりさらに5000万年前のジュラ紀に出現したとする遺伝学からの証拠が、古生物学者にも受け入れやすくなったのである。
しかしながら、仮に哺乳類の多様化がK-T境界絶滅の前に進行していたとしても、最も目立つ変化である大型化は恐竜が絶滅した直後に起きた。その後の27万年間で、哺乳類は多様化するとともに体も大きくなっていったのである。
ただし、本当の意味で大型と呼べる哺乳動物が出現したのは、約5500万年前になってからであった。当時、地球の温度が急速に上昇し、世界中に森林が広がり、南北両方の極地付近にも樹木が生い茂った。これは、哺乳類の多様性の大幅な拡大を後押しした可能性がある。
暁新世の陸の世界
『暁新世』は、K-T境界の大量絶滅があったからこそ訪れた時代である。
この大量絶滅の原因と結果については疑いようがない。その後の世界は、多方面で全く新しいものへと変化した。
陸上では恐竜の支配があまりにも長かったため、絶滅を生き延びた生物による新たな生態系の構築が急務となった。その上、数多くの陸上動物が突如として消滅したことで、進化の歯車が動き出し、新種が次々と生まれるようになり、動物の多様性はカンブリア爆発にも匹敵する勢いで増加した。陸上では哺乳類が顕著な勝者となったことは間違いないが、鳥類も返り咲き、しばらくの間、陸生哺乳類と様々な資源を巡って競い合っていた。
巨大な隕石が落ちたのは、低酸素と温暖化が長く続いた中生代の時代であった。この隕石の落下が気候に及ぼした影響は大きく、何千年もの間、生態系全体に余波が広がった。まず、すでにわずかに寒冷化していた世界で、陸地と海洋の気候がますます不安定になった。生物相の変化も同様に壊滅的であった。恐竜がいなくなった結果、森林が密林化した。現代でも、象が歩き回ったり植物を食べたりすることで森林の密林化を防いでいる ように、象よりも遥かに体が大きかった恐竜もまた、植生パターンを左右する存在であった。その恐竜が突然姿を消したために、森は鬱蒼となった。
K-T境界絶滅から700万年以上が経過した暁新世後期には、気候はすでに世界的に安定していた。地球はゆっくりと温暖化しており、全世界の温度が上昇した。酸素同位体比の調査から、赤道付近の海洋表層水の温度は20℃を超え、場所によっては26℃に達していたことがわかった。つまり、現代の赤道付近の海水温に近かったのである。現代と大きく異なるのは高緯度地域である。北極や南極付近の表層水の温度は、現在はほぼ氷点であるのに対し、当時は10~12℃であった。したがって、赤道と極地の水温差は10~15℃程度と、現在の半分ほどしかなかった。しかし、こうした違いにもかかわらず、海流のパターンは現在と非常に似ていた。特に重要なことは、最終的に海底に沈み込む高酸素の水塊が、現代と同じように高緯度地域で発生していたという事実である。
約6500万年前にK-T境界の大量絶滅が発生した後、生存した哺乳類が植生パターンに影響を及ぼすほどの大型化を遂げるまでには何百万年もの時間が必要であった。恐竜の死骸が腐敗し、悪臭を放つ世界で、ネズミほどの小型哺乳動物が巣穴から這い出す光景は、多くの絵画で描かれている。何か月間かの間、死肉を食べる哺乳動物にとっては至福の時期であったと考えられる。しかし、骨がすぐになくなり、それさえも短期間で朽ち果てるか土に埋もれると、全ての哺乳類は、以前とは全く異なる食物網の中で生きることを余儀なくされた。暁新世初期には草がまだ出現していなかったため、植物食動物は草ではなく、木の葉や果実を食べていた。しかし、木の葉を主食とする生物は少数であったと見られる。暁新世の哺乳類の歯の形状から推測するに、固い葉よりも昆虫や果実、柔らかい若枝を常食とする生物が多かったと思われる。また、根や塊茎を食べる生物も存在した可能性もある。
木の葉に適した歯の形状が出現したのは、暁新世後期のことであった。しかしひとたび進化が始まると、新しい種類の哺乳類が次々に誕生し、哺乳類の大型化も加速した。そしてK-T境界の大量絶滅からわずか900万年後、生物が生息する環境は再び危機に見舞われることとなった。
暁新世-始新世環境温暖化極大イベント(PETM)
地球は新生代初期(古第三紀の初期)の時点で、記録されているだけでも「少なくとも9回の大量絶滅」に見舞われている。
- 最初は古原生代の22-23億年前に起こった『大酸化事変とそれが引き起こしたスノ―ボールアース現象』による絶滅(<物語り その7>「大酸化事変とスノーボールアース」の節参照のこと)。
- 二度目はそれから10億年以上のちの『クライオジェニアン紀』(<物語り その8>「生命とスノーボールアース現象」の節参照のこと)。
- その後は年代順に『エディアカラ紀後期』(<物語り その8>「世界に点在するエディアカラ生物群」の節参照のこと)、『カンブリア紀後期』(<物語り その9>「カンブリア紀の終焉 - 顕生代初の大量絶滅」の節参照のこと)、
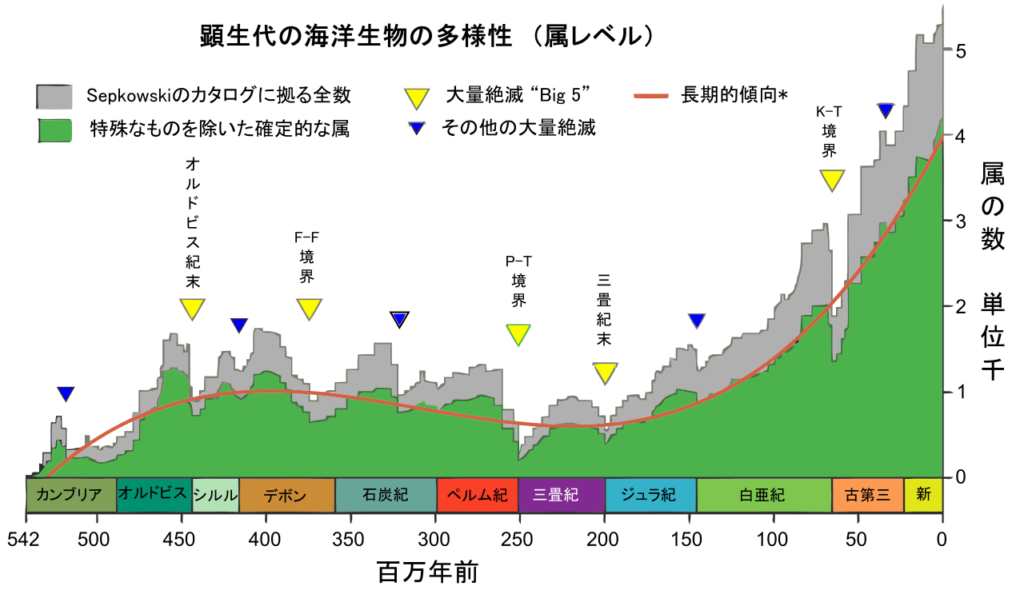
- そして「ビッグファイブ」と呼ばれる五度の大量絶滅『オルドビス紀後期』、『デボン紀後期』、『ペルム紀後期』、『三畳紀後期』、そして『白亜紀後期』と続く(図229参照、黄色逆三角にて表示)。
原因は多岐にわたり、酸素濃度の急激な上昇や酸素欠乏、捕食者の出現や硫化水素の放出に伴う無酸素状態、果ては隕石の衝突までが含まれる。
(注182)顕生代542百万年を通した海洋生物の属レベル多様性の推移を分かりやすく示した図である。Sepkoski, J. 著「化石海洋動物属の一覧表」(2002)に記録された36380属に基づいている。なお、緑のグラフは、当該属の出現と絶滅の時期が明確に特定されているものだけをカウントした場合である。Raup and Sepkoski (1982)により命名された”Big Five”は黄色三角、それ以外の絶滅イベントは青三角にて示した。
しかし、恐竜が絶滅したわずか900万年後の暁新世末に、新たな脅威が登場した。それが『メタン』であり、これが記録されている中でも特に『急激な地球温暖化』を引き起こした。
- この現象は『暁新世-始新世境界温暖化極大イベント(Palaeocene-Eocene Thermal Maximum; PETM)』と呼ばれる。
暁新世と始新世の境界付近の地層では、火山灰が全世界で著しく増加している。火山灰はきわめて微細な物質なので、塵と同様に大気から海底へと進んでそこに堆積するが、発生の原因は火山の噴火である。
- 火山灰が増えているとすれば、5800万年前~5600万年前に地球全体で火山活動がにわかに活発化したとしか考えられない。
- 世界中の様々な地点でさらなる調査が行われた結果、これが一つの海盆だけに起きた異常現象ではなく、地球規模のイベントだったことが確認されている。
暁新世後期には、熱帯地方の気温は以前とほぼ同じであったが、極地方は著しく温暖化した。赤道と極地の海水温差は、暁新世においては17℃(現在はさらに大きく22℃)であったが、始新世前期にはその差はわずか6℃に縮小した。極地の温度が上昇するにつれて、熱帯-極地間の熱交換は鈍化し、嵐の頻度と強度も減少した。
世界は再び、穏やかな気候の非常に暑い場所となり、『温室効果大量絶滅の環境』と化した。
暁新世-始新世の境界層をまたいで採取した二つのコアからも、炭素の同位体に大きな変化が確認されている。
- 同位体比(δ13C=[(13C/12C)試料/(13C/12C)標準-1]x1000(‰)、注57<物語り その7>を参照のこと)に短期間ながら負の変動が見られたのであった。これは、植物量の減少時に観察され、大量絶滅を明確に示す特徴である(注183)。
(注183)光合成の際、生物は軽い炭素(12C)をより多く固定するので、その結果、海水には重い炭素(13C)が多くなる。逆に、生物量が減少すると、軽い炭素(12C)の固定量が減少するため、海水には重い炭素(13C)が相対的に少なくなる。そのため、植物量が減少すると、海水の炭素の同位体比(13C/12C→δ13C)が変化、減少するのである。
古生物学者の中には、その地域の底生生物(水域の生物のうち、水底を這い回ったり、穴を開けたり、また、水底や壁面に付着するような生物)の生存状況に目を向ける者もいた。彼らがとくに注目したのもやはり「底生有孔虫」で、海底でも壊滅的な大量絶滅が起きた証拠が見つかった。こうした事実が1990年代の初めに明らかにされると、ほどなくして日本の古生物学者、海保邦夫が別の研究結果を発表した。それによると、底生生物の運命を決したのは深海の温度上昇ではなく、海底の酸素濃度の低下であったとの結論が導かれた。この仮説は直感的にも説得力があり、理由としては温かい水が栄養豊富である一方、酸素不足になりやすいからである。
海洋の深層が温暖化し、酸素濃度が低下すると、その影響は表層の水温まで及んだ。その原因となったのは何だったのだろうか。海底全体の温暖化を引き起こしたのは、熱帯の温かい表層水であったと考えるのが適切であるだろう。表層では蒸発により海水の塩分濃度が上昇し、その結果、密度が大きくなる。この温かくて塩分濃度が高い海水が沈んで海底に沿って運ばれ、高緯度地方の低温地域まで遠く移動するのである。
- 通常なら、冷たく酸素濃度の高い表層水が深海底に向けて流れていくのだが、暁新世末の海洋ではこの流れが機能していなかったのである。
- 『深層の熱塩循環システム』(海水は主にこれにより混ざり合っている)が、現在の海洋に見られる流れと正反対なのである。
- 最初に影響を受けたのは酸素を必要とする微小な生物、つまり深海の『底生有孔虫』(<物語り その15>図222を参照)であり、その多くが約40万年という比較的短期間で急激に絶滅したのである。
しかしながら、大量絶滅を確かに指摘するためには、海洋だけでなく陸上の生物も影響を受けたことを明らかにしなければならない。そのため、陸上での変化を探る調査が始まった。古生物学者は、深海での絶滅の発見に刺激を受け、暁新世末に陸上でも絶滅があったのかを確認するため、当時の陸上生物の化石記録を再検討し、新たな化石も収集した。その結果、哺乳類間で大きな変革が起きていたことが明らかになり、年代を精確に調査した結果、陸上と海洋の絶滅は同時期の出来事であったことが確認された。
- PETMによる温室効果が海洋生物に引き起こした大きな変化は、陸上でも発生していたのである。
- 陸上生物の化石記録を見る限り、この変化が現在の哺乳類動物相の始まりを示していると思われる。
暁新世の後半にはすでに多様な哺乳類が存在していた(明らかに異なる30の科が化石から確認されている)、多くは小型であり、現存しない分類群のものも含まれていた。例えば、『齧歯類(げっしるい)』(注184)に似た小型動物の生き残りや、様々な『有袋類』、アライグマに似た『有蹄類』などである。真性の『食虫動物』や最初の『霊長類』(食虫動物と同様にまだ小型)も存在していた。しかし、暁新世後期に入ると、いくらか大型の生物が出現し、中には非常に奇妙な生物もいたのである。
(注184)鑿 (のみ) 形をした一対の門歯は絶えず伸びつづけるので、常に物をかじってすり減らす。犬歯はない。哺乳類では最も種類が多く、リス・ネズミ・ヤマアラシの3亜目に大別され、約1600種が知られる。
しかし、暁新世後期に入ると、いくらか大型の生物が出現し、中には非常に奇妙な生物もいたのである。
例えば、『汎歯目』というグループには、イヌからバイソンくらいの大きさの草食動物がいて、一部はカバのように半水生で生活する種や樹上生活をする種に分化し、さらに大型化して森の地面を四肢で歩きまわる種も存在した。一般的には体が矮小で足も短いため、少なくとも現代の草食動物と比べると歩行はかなり不格好であったと思われる。汎歯目は大型であったが、暁新世末にはさらに大きな草食動物が出現していた。それが巨大な『恐角目』で、特大のサイのような動物で、頭部に奇妙なこぶや角を一式備えているところまでサイに似ていたのである。


暁新世から始新世への移行を示す地層においては、一時的に種の数が減少し、その後時間を経て徐々に新種が出現する過程が観察される。これら地層から見つかる骨の大部分は、我々が以前から知っている種類に所属するものである。例として、最初の偶蹄類(注185)が出現する。そしてすぐに、より現代的な肉食獣が現れ、新しい植物食動物を捕食する形で進化していく。これらの肉食獣は、現生肉食動物とも系統関係にある。
(注185)偶蹄類は指行性(かかとを浮かせた状態で指先だけを地面につけて歩く歩行形態)であり、親指が退化して指数が偶数となる。体重は主に第3、第4の指にかかる。多数が草食で、反芻胃を持つ種も存在する。現生の偶蹄類は、イノシシ、ペッカリー、カバ、ラクダ、マメジカ、シカ、キリン、プロングホーン、ウシの9科に分類され、約150種が知られている。
これらの変化は全て、世界の気候を変えた出来事の結果である。過去の大量絶滅から我々が学べることは、大規模な絶滅によって新たな形態の生物が出現する機会が提供されたという事実である。暁新世の終わりにも同様の出来事が起こっている。
- まず、PETMは私たち人類が無関心でいることができない重大な出来事であると指摘している。
- その理由は、PETMの時期に12兆~15兆トンの炭素が大気中に放出されたが、これは人間が長年にわたり工業生産やエネルギー利用のために排出してきた炭素の量とほぼ同等であるからだ。
- 加えて、PETMでは温室効果ガスの増加により気温が上昇し、当時の世界は今より5~9℃も温暖であった。PETM自体はおよそ1万年にわたって続いた。
PETM期間中に生息していた植物群は、その前後の時代のものとは大きく異なっている。例えばマツ類などの裸子植物はすべて姿を消していた。代わりに存在していたのは、PETM以前はより低緯度、つまりより温暖な環境で生きていた植物群であった。これはスミソニアン博物館の古植物学者スコット・ウィングの発見に基づく。
PETMが終わると、以前の植物群が再び現れた。昆虫も同様で、1万年間の激しい環境変化の後に、以前生息していた種が再び現れた。しかし、哺乳類については同じことは言えない。
この温暖化現象は北米の哺乳動物相を一変させたのである。
さらに重要な点が一つある。
もし当時、現在の地球のような大規模な氷床が存在していたら、それは間違いなく急速に溶け、海面上昇を引き起こしていたであろう。これこそが人間による温暖化の最も危険な側面である。現在、我々人類は南極大陸やグリーンランドの氷を溶かしてしまっている。今後数世紀で、現在農地として使用されている広大な土地が水没する可能性がある。海面上昇率が最も高いと現在確認されている地域は、中国南部の海岸地帯である。この地域は世界でも人口が密集しており、海抜ゼロメートルの地点には広大な水田が広がっている。
寒冷化する新生代の草原と哺乳類
『始新世』から『中新世』の初めにかけて、世界はゆっくりと寒冷化していった。まだ最初の始新世のうちはごくわずかな変化にすぎず、現に当時は地球規模で熱帯林が広がっていて、現在の北極圏内にワニがすむほどだった。しかし、
『漸新世』に入ると冷却が加速し、気候の性質が大きく変わり、それまでほぼ均質だった世界の気候には季節性が生まれた。同時に、大規模な『大陸氷床』が南極大陸とおそらくグリーンランドでも形成され始めた。この氷床の拡大は急激な海面低下を引き起こし、高緯度地方では森林が草原やサバンナに置き換わった。しかし、それだけではない。
- 大気中にも変化が生じ、これが生命の歴史に多大な影響を及ぼすこととなった。
植物には二酸化炭素が必要であるが、二酸化炭素の濃度は数十億年にわたって増減を繰り返している。しかも、その背後には一つの長期的な傾向がある。それは
- 細かい変動はあるものの、長期的に見ると二酸化炭素の量は減少しているという事実である。
- 二酸化炭素が長期にわたって減少すると、地球は徐々に冷却される。特に過去4000万年はそうであった。
しかし、新生代の植物の進化に影響を与えたのは気温の変化だけではない。
- もっと重要なのは、二酸化炭素の減少に呼応して、「より効率的な光合成」の方法が生まれたことかもしれない。これは『C4型光合成』と称されている。
- 多くの植物は従来の『C3型光合成』を放棄し、「C4型」を採用するようになったのである(注186)。
- それどころか、C4型を使う植物が増えたことから、C4型光合成は大変な勢いで重要性を増してきている。
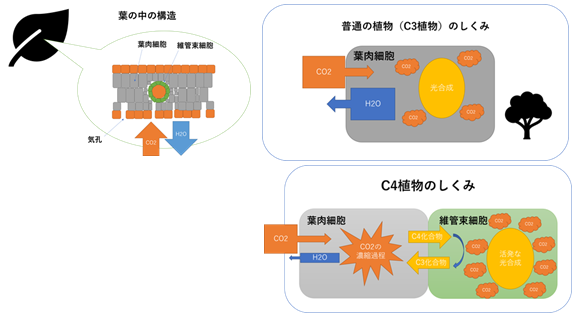
(注186)C4型光合成とは、光合成の過程で一般のCO2還元回路である『カルビン・ベンソン回路』の他にCO2濃縮のための「C4経路」を備える光合成の形態である。「C4経路」という名前は、CO2固定における初期産物である「オキサロ酢酸」が「C4化合物」(炭素が4つの化合物、すなわちコハク酸、フマル酸、林檎酸、オキサロ酢酸など)であることに由来している。C4型光合成を行なう植物は『C4植物』と称され、「維管束鞘細胞」にも発達した葉緑体が存在するのが特徴である。これに対して「カルビン・ベンソン回路」しか持たない植物は『C3植物』と称される。
普通の植物(C3植物)は気孔から吸収したCO2をそのまま利用して葉肉細胞でC3化合物を生成するが、C4植物は気孔をほとんど開かずに水蒸気が逃げるのを抑えつつ、体内の二酸化炭素の濃度を高めて効率よく光合成を行う。したがって、C4植物は普通の植物よりも少ない二酸化炭素でもより活発な光合成を行うことが可能になるとともに、同時に水蒸気が逃げるのも抑えることができる。
C3型光合成を行なう植物とC4型の間には、炭素同位体比における違いが残される。これは、生体組織を特定する質量分析計を使用して植物組織を分析することで測定可能である。さらに、その植物を摂食した動物も必ずその痕跡を留める。したがって、化石を調査することで、特定の種の植物食動物がC3植物を食べていたか、C4植物を食べていたか(あるいはその両方だったか)を明らかにすることが可能である。
C4植物が初めて出現した時期を示す証拠は、二つの方向から得られる。すなわち、ゲノム比較の分子時計による結果(おそらく最も早くて2500万年前)と、化石記録から得られる結果(およそ1200万年前~1300万年前)である。しかしながら、これら二つの方法ではまったく異なる答えしか得られていない。C4型光合成は、革新的な進化が広範に拡散する種類のものではなく、実際には40以上の異なる系統の植物で個別に進化した可能性があるのである。
こうして生まれたC4植物は、火や乾燥に強く、高温かつ乾燥した気候に適応している。
- C4植物の中でも特に重要なのは『イネ科植物』である。それは、大型の哺乳類や様々な鳥類(例えば都市部でも水辺の芝生でよく見かけるカモなど)を含む多種多様な植物食動物の食生活の中心になっているからである。
二酸化炭素が特にここ2000万年間で減少していることが、C4植物の草原が拡大するのを大いに後押ししている。
- ほとんどのイネ科植物は森林の地面では生存できない。日陰が多く、比較的涼しい環境は生育に適さないからである。しかし、森林が破壊されると土地が解放され、イネ科植物にとっては遙かに適した生息環境が形成される。
長らく主流だった考え方は、二酸化炭素の長期的な減少がC4植物の進化を促し、イネ科植物が優位に立つことを助長したというものであったが、近年では新たな見解が提唱されている。
- それは、「森林被覆率」の変化が二酸化炭素の減少と同様に、あるいはそれ以上に重要だったという見解である。
では、森林の急激な減少を引き起こした原因は何であったのか。その答えは、「火事」である可能性が高い。
地球は植物の覆う惑星であるが、その中で『森林火災』の影響は過少評価されている。火災は当然ながら酸素濃度の影響を受ける。酸素濃度が高かった時代、とくに3億2000万年前~3億年ほど前の石炭紀には、森林火災は頻繁に発生していたと推測される。この時期に地球を宇宙から観察すると、煙が大気を充満し、視界が暗く霞んでいた可能性が高い。地球全体が靄に包まれ、すっきり晴れる日などめったになかったかもしれない。そのような煙が大陸の大部分を覆っていたとしたら、地球の気温に極めて重大な影響を与えていたと考えられる。なぜなら、森林火災の煙は上空から見ると白っぽい色をしており、靄や煙がないときより多くの太陽光を宇宙に反射するため、「アルベド」(惑星が太陽光を反射する度合い)が変化し、結果的に気温が下降すると考えられるからである。
これら全てが相互に影響しながら、その後の地球の気候はもちろん、生命の歴史にも大きな変化を引き起こした。
- 酸素濃度が上昇し、石炭紀全体の三割以上の期間で高い状態が持続したために、森林火災が増えた。
- 前述の通り、地球の気温は下降し、これに続くさまざまな出来事が積み重なって結果として極地が氷河に覆われた。凍結期間の長さは一部のスノーボールアース現象の時期とほぼ匹敵したであろう。この氷河期は5000万年以上続いた可能性がある。
時期を同じくして、地球史上最も重要な出来事がいくつか起きている。
- 一つは、動物が陸上を支配し始めたことである。また、それまでの進化の過程で(当時としては)高等とされる陸上植物が新たに登場し、それまで生息できなかった大陸の高地に群生できるようになった。
- その後、脊椎動物の中でも特に重要なグループが登場した。これには、最初期の爬虫類と、その直後に出現した哺乳類の祖先などが含まれる。
しかし、火事には別の側面も存在し、それが植物の進化史、そして広くは生命の進化全般に影響を及ぼしたと考えられる。
アマゾン川流域の火災に関する新たな研究により、野火が気候に大きな影響を及ぼすこと、そしてそれが熱帯限定の現象ではないことが証明された。イギリスの古気候学者デイヴィッド・ビアリングは、著書『植物が出現し、気候を変えた』の中で、1988年の火災による煙が北米の一部で雲の形成を妨げ、それが降雨パターンを変化させた可能性を指摘した。1988年の四月には深刻な干ばつが発生し、20世紀でも最悪の乾燥が何か月も続いた。この干ばつの前には、未曾有の規模の森林火災が発生していた。同じく大規模な森林火災は、1988年の7月にも北米で発生しており、この年のイエローストーン国立公園一帯が広範にわたり焼失した。
ビアリングは、C4植物の草原が拡大したメカニズムに関して新たな視点を提供した。彼は、
- 森林火災にも正のフィードバックが働き、さらなる森林火災を引き起こすとの観点を示した。火事が気候を変変化させ、干ばつが起きる頻度が高まる。
- そのせいで燃えやすい地域が拡大し、火事の被害がさらに増大する。そして、それによって気候が更に変化し、この循環が繰り返される。火事がさらなる火事を生じるという過程により、結果的にC4植物が分布を広げたという見解である。
図表
図226 モルガヌコドンの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「モルガヌコドン目」(https://ja.wikipedia.org/wiki/%E3%83%A2%E3%83%AB%E3%82%AC%E3%83%8C%E3%82%B3%E3%83%89%E3%83%B3%E7%9B%AE#:~:text=%E3%83%A2%E3%83%AB%E3%82%AC%E3%83%8C%E3%82%B3%E3%83%89%E3%83%B3%E7%9B%AE%20(Morganucodonta)%20%E3%81%AF%E3%80%81,%E3%81%9F%E7%B5%B6%E6%BB%85%E5%8B%95%E7%89%A9%E3%81%A7%E3%81%82%E3%82%8B%E3%80%82)
図227 臼歯の咬頭
おのざき歯科医院
(https://www.onozakishika.com/15_kamiawase.html)
図228 エオマイアの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「エオマイア」
(https://ja.wikipedia.org/wiki/%E3%82%A8%E3%82%AA%E3%83%9E%E3%82%A4%E3%82%A2)
図229 顕生代における海洋生物多様性(属レベル)の推移
Wikimedia Commons, the free media repository “File:Phanerozoic Biodiversity.png”
(https://commons.wikimedia.org/wiki/File:Phanerozoic_Biodiversity.png)
図230 パントラムブダ(原始的な汎歯目動物)の想像図
フリー百科事典『ウィキペディア(Wikipedia)』「汎歯目」
(https://ja.wikipedia.org/wiki/%E6%B1%8E%E6%AD%AF%E7%9B%AE)
図231 ウインタテリウム(恐角目動物)の想像図
フリー百科事典『ウィキペディア(Wikipedia)』「ウインタテリウム」
(https://ja.wikipedia.org/wiki/%E3%82%A6%E3%82%A4%E3%83%B3%E3%82%BF%E3%83%86%E3%83%AA%E3%82%A6%E3%83%A0)
図232 C3型・C4型光合成のしくみ
京都の造園会社・大幸造園のスタッフブログ・庭園日誌
(http://daiko-zoen.blog.jp/archives/76167064.html)

{kind=link}