
第10章 温室化した海【2億 ~ 6500万年前】
大気と海は密接に相互作用を持つため、大気の状態は海洋の化学的・物理的環境に重大な影響を及ぼす。大気の温度、極地と赤道の温度差、海水の化学的組成(溶存酸素量も含む)はすべて、海洋の状態やそこに生息する生物の種類を左右する要素である。
重要な点は、温水は冷水より酸素含有量が少ないという事実である。白亜紀末の500万年を除き、中生代には地球の大気が極地から赤道まで高温多湿であった(注165)。この高温は、海洋の酸素濃度を現在よりも低くする効果があった。さらに、大気中の酸素濃度が低かった(<物語り その10>図114参照)ため、中生代の海洋環境は現在と大きく異なり、生命にとって過酷であった。そこで生息していた生物は、この低酸素状態に対応するための様々な進化を遂げていた。
(注165)古生代の終わり頃、大陸はパンゲアと呼ばれる超大陸を形成し、一か所に集まっていた。しかし、中生代(約2億5200万年前から始まり、三畳紀・ジュラ紀・白亜紀と進む)に入ると、パンゲア超大陸は分裂し始め、火山活動が活発化した(<物語り その14>図198参照)。この火山活動の結果として放出された大量の二酸化炭素は温室効果を引き起こし、中生代全体が温暖化した。特に白亜紀は、地球史上で最も温暖な時期であったと考えられている。熱帯は拡大し(ペルム紀には熱帯の範囲は北緯20度から南緯20度だったが,三畳紀には北緯23度から南緯23度に拡大した)、年平均気温は25から30℃であった。地球全体の平均気温は現在より10℃程度高く、北極や南極には氷が存在せず、海水面は現在より約200mも高かった時期があったと推測されている。
中生代の世界は現代とは大きく異なるものの、ある一点では類似している。それは、空が動きと生命で満ちていたという点である。現代では、昆虫から鳥、コウモリまで、様々な飛行生物が多数生息している。
中生代にも、現代とは大きく異なる二つのグループが存在していた。一つは爬虫類の『翼竜』と小型の『翼手竜』、もう一つは形状が現代の一般的な鳥とは異なる鳥類である。その鳥類には、歯があるものやないもの、翼のあるものやないものなど、多様性が見られた。


海の生物相について考えると、白亜紀の時期には多くの海洋地域に「ラグーン(潟)」が広がっていた。ラグーンとは、礁などが壁となり外海と内海を隔てた場所で形成される。ラグーンは外海より水温が高く、酸素も少ないのが一般的である。白亜紀のラグーンの浅瀬には、二枚貝や巻貝が生息していた。これらの生物は、現代の熱帯ラグーンや海岸近くの環境に生息するものと類似しており、分類学上の同じグループ(「属」など)に分類されるものも多い。例えば、ミルガイ、ツノガイ、カキ、イタヤガイ、イガイ、タカラガイ、イモガイ、ホラガイ、コンクガイ、エゾバイ、ウニ(岩などの表面で生活する一般的な「球状」のウニと、今日のスカシカシパンやタコノマクラのように穴を掘る「特殊な」ウニの両方)などが存在していた。また、トゲのあるロブスターやカニも見られたであろう。
全体的に見ると、白亜紀後期(9000万年前~6500万年前)の浅瀬にはすでに「現代的」な動物相が確立していたと考えられる。その頃から、白亜紀末の大規模な大量絶滅イベントに向けて時間は刻まれていた。それにもかかわらず、その影響は最終的にそれほど大きなものではなく、生物相は比較的安定した状態を保つことができたと思われる。
現在の海と同様に、水深が深くなると生物の種類が変化する。深部へと進むと、堆積物の粒子が微細化するため、浅瀬で生活する粗い砂に適応した種よりも、微細な砂に適応した生物が主となる。多種多様な生物が砂に潜んで生活していたと推測される。現存する二枚貝と同様の種も多く存在し、穴を掘り、そこで生活する様々な種もいたと推定される。堆積物に身を隠すという生存戦略が主流であった。白亜紀後期には多種多様な捕食者が存在し、軟体動物の殻を壊す、あるいは穴を開ける能力を持って進化していた。ラグーンの浅瀬には、現代のサンゴに相当する造礁生物が活動し、硬い石灰岩を形成していた。この石灰岩の塊は小さな馬蹄形の礁を形成し、卓越風が吹く方向に礁のアーチ部分が向かうように進行した。これは現代の礁形成プロセスと同様である。
海岸から離れた地点には、海面まで達する大きさの『堡礁(バリアリーフ)』が存在していたと推測される。大陸棚が終わる地点には石灰岩の巨大な壁が数百キロから数千キロにわたって延び、大きな島や大陸を縁取っていたと考えられる。その壁の内側と外側には、硬骨魚類や軟骨魚類(サメ、ガンギエイ、アカエイ)など多種多様な魚類が生息していたとされる。このバリアリーフは形状的に現代のオーストラリアのグレート・バリアリーフに似ていたと考えられる。しかし、その形成に関与する生物群は現代と大きく異なっていた。
- 現代の礁にはサンゴが生息しているが、白亜紀の礁の骨格を形成していた主体はサンゴとは全く異なっていた。
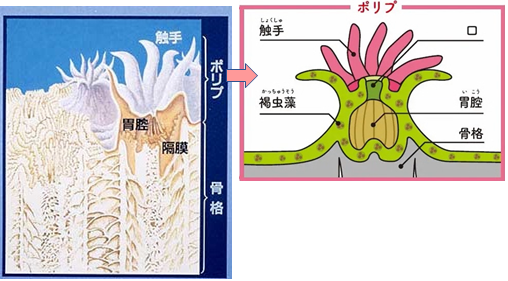
『礁』とは波に耐える立体的な構造で、オルドビス紀以来、生物が集まる重要な場所として機能してきた。すべての礁は、骨格を形成する生物とそれを結合する要素によって構成されており、現代のサンゴ礁もこの基本的な構成要素は変わらない。それはまるでレンガ造りの家のようで、サンゴの「レンガ」、被覆性の藻、平たいサンゴ、そして炭酸塩粒子の「モルタル」から成り立っている。
何百年もの時を経て、より大きくてどっしりしたサンゴが既存の礁の表面に付着して積み重なり、太陽光に向かって上へ上へと成長する。周囲のサンゴより速く成育できるかどうかは、生き残るか否かを決定する重要な問題である。サンゴは頭上を覆われないようにするために競争する。さもないと、生命を育む太陽光と、開けた水域を利用できなくなるからである。太陽光は、ポリプ内に共生している何百万もの植物プランクトンのために必要であり、開けた水域は肉食性のポリプの食物をもたらす。共生している植物プランクトンは「褐虫藻」(渦鞭毛藻類の一種)と呼ばれ、サンゴが巨大な骨格を築くのを助ける代わりにサンゴから栄養をもらい、捕食者から守ってもらっている。微小なサンゴの幼生は海を漂い、無生物の硬い基盤を見つけるとそこに固着し、やがて海面に向かって成長する。幼生は単体のポリプから始まり、運が良ければ数百、数千のポリプが集まる巨大な群体となり、何百年、何千年と生き続けることができる。巨大な石灰質の骨格は重さ数千トンにも達することがあり、現在確認されている中には数千年以上生存する群体も存在する。しかし、どれだけ巨大な群体であっても、最終的にはその生命を終え、その骨格は崩壊し、次世代のサンゴの基盤となる。
白亜紀の温暖化した海においても、同様のプロセスが進行し、最終的な形状も現代のサンゴ礁と変わらない。しかし、その礁を構成する生物が現代のサンゴとは異なる。
- その主役は二枚貝で、『アツバニマイガイ(厚歯二枚貝)』(注166)と呼ばれる種であった。
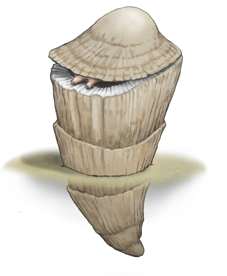
(注166)ジュラ紀に出現し白亜紀末に絶滅した中生代特有の二枚貝である。特に白亜紀中期以降に繁栄し、世界各地の低緯度〜中緯度海域で大規模な礁を形成した。厚歯二枚貝は白亜系の石灰岩の中に多産するため、ヨーロッパでは建築石材に含まれる身近な化石として知られている。厚歯二枚貝の多くは現生のカキやシャコガイのように、片方の貝殻で海底に固着した生活を送っていた。この生態のためか、非常に奇妙な形状の殻を持つものが多い。また、厚歯二枚貝としては比較的大型で厚手の殻を持ち、殻の内部や表面に複雑な構造を発達させた。二枚貝の分類は一般的に貝殻の蝶番部分にある歯(鉸歯)の形状が決め手となる。厚歯二枚貝では厚手の鉸歯が非常に大きく発達する。そのため、「厚い歯の二枚貝」と呼ばれるようになった。これが和名の『厚歯二枚貝』の由来である。厚歯二枚貝はこの蝶番部分がきわめて頑丈にできていることから、立体的で多様な殻の形状を獲得することができたと考えられている。
もっとも、現生の二枚貝と比べると、その外観は全く異なっていた。特異な形状を持つものが多く、一部は垂直に立てたゴミバケツに似ており、筒状の殻の上には開閉可能な蓋さえ備わっていた。中には現代の熱帯に生息するオオシャコガイに近い大きさのものも存在した。しかし、独立して生活するシャコガイとは違い、現存するイガイのように群生する特性を持ち、底部を覆い尽くしたり、個体同士が互いに重なることもあった。
大きな「筒」の部分はアツバニマイガイの下側の殻に相当する。これらが縦に立ち並び、隙間なく配置されることで、全体として強固な石畳のような形状を形成した。石畳の「舗石」に相当する個体は高さ約30~60センチ、直径が時折30センチにもなる円錐形で(下部が細くなっている)、それぞれの上部から鮮やかな色の軟体部を光に向けて伸ばしていた。サンゴと同様に、小さな単細胞の藻と共生しており、その藻の光合成に光が必要であった。見返りとして、藻はアツバニマイガイに多量の酸素を供給し、その組織から二酸化炭素や排泄物を取り除いていた。
しかし、現代のサンゴが大きくなるには何百年も必要とするのに対し、アツバニマイガイの成長速度は極めて速かった。浮遊プランクトンの状態から、浅海の海底に沈み(藻を共生させるために光が必要だったと考えられる)、1年未満で炭酸塩の厚い外殻を形成し、成体の大きさに達した。一度生まれると短期間で成長し、ほとんどがすぐに死亡した。その理由は、同じ種の個体が硬い殻の上に侵入し、勝手に「領地」(生きてはいるが動けない)を占拠し、窒息させてしまうからであった。高さと幅が2メートル近くに達する群体を形成するためには、サンゴの骨格であれば約100年を必要とするところが、アツバニマイガイではせいぜい5年しか必要としなかった。
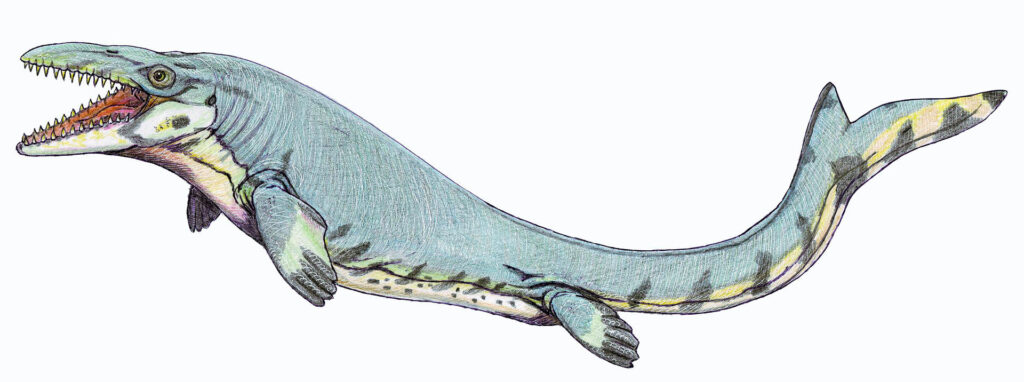
アツバニマイガイの礁の海側では、水深が急に深くなる。礁の外側に広がる中生代の広大な海洋には、今では絶滅した多種多様な生物が生息していた。表層水域には大型のサメや巨大な海生爬虫類が泳いでいたと考えられる。海生爬虫類には、首が長い(または短い)プレシオサウルスや、ワニに似たモササウルスがいた。これらはおそらく現代のアザラシのように、捕食のために潜水する一方で、呼吸のために海面に浮上する必要があった。
ただし、これらは任意のアザラシよりもはるかに大きく、休息時や出産(あるいは産卵)時に水から出る生物と比較しても大型であった。
当時の温暖化した海では、深層の状態も現代の一般的な海洋とは異なっていた。現在では黒海が唯一、当時の深層・中層水域の状態に近似している。そこは温暖で溶存酸素が非常に少なく、魚類すらほとんど生息できない環境であった。海底は黒海と同じく黒い泥で覆われていた。この泥には細かな微粒子状の黒い有機物が大量に含まれていた。こうした深海では海水の酸素濃度が極端に低く、そのために有機物を通常の方式で分解することが難しく、酸素が豊富な海底に比べて分解率ははるかに低かった。
この泥状の海底堆積物の表層10センチくらいには、通常とは全く異なる微生物群が生息していた。それらは硫黄で生きる微生物であり、その特異な副産物として放出されたのが硫化水素やメタンといった化合物であった。
中生代の海では、通常の動物が生きていけるだけの酸素が海底に存在する地域は極めて限られていたと考えられる。しかし、そのような低酸素環境に適応した二つの軟体動物が現れた。
- 一つは海底に生息する『二枚貝類』の軟体動物。
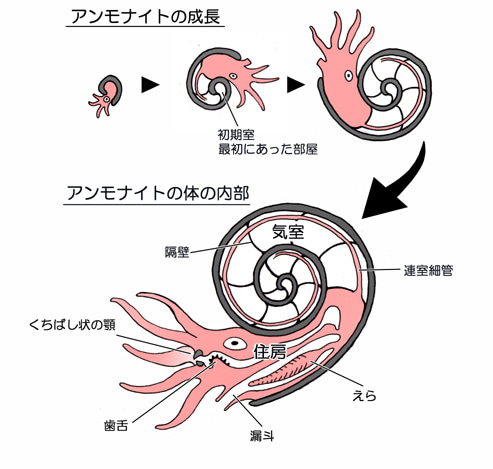
- もう一つは多種多様な頭足類の軟体動物、アンモナイトである。アンモナイト類は水中を生息域とし、摂食は海底で行われた。
ここで紹介する白亜紀のアンモナイトが所属するグループは、ジュラ紀の始まり頃に出現した。その時代の岩石にその存在が突如として記録されていることから、三畳紀-ジュラ紀の境界の壊滅的な大量絶滅(<物語り その10>図113参照のこと)の後で、新種の動物が登場する道が開かれたと推測される。その一つが『新しいデザインのアンモナイト』であった。
このアンモナイトの最後のグループはジュラ紀最古の地層から始まり、まさに本章の主題である温暖化した海の地層に至るまで続く。これらは生命の歴史において大きな意味を持つだけでなく、化石から年代を割り出す作業、それに続く地質学そのものにとっても極めて重要である。三畳紀の末に海成層の上にジュラ紀の地層が積み重なった場所は、世界中に数多く存在する。そうした地層が露出している場所では、歩きながら時代を追体験することが可能で、地層が連続していれば、三畳紀後期からジュラ紀前期にかけての劇的な変化を直接目の当たりにすることができる。
この時代の岩石には、『ビッグファイブ』と称される大量絶滅の一つ、『三畳紀末の大絶滅』の証拠が保存されている。これは膨大な数の生物種が絶滅したという、あまり肯定的とは言えない事象の記録である。三畳紀後期の地層を調査すると、最初に『ハロビア』と呼ばれる平らな二枚貝の化石が密集した層が見つかる。その後、新しい時代の地層へと移ると、同じ二枚貝である『モノチス』が大量に発見される。しかしながら、そこから地層がわずか数メートル進むと二枚貝は姿を消し、その後は化石が存在しない岩石が続く。これが三畳紀最後の地層で、恐らく300万年間にわたるものであった。


この化石がほとんど見られない厚い地層の後に、突如として新しい生物群が現れる。それは『アンモナイト』である。
- 三畳紀後期の岩石にもアンモナイトの化石は存在するが、その数は限られている。しかし、ジュラ紀初期のアンモナイトは、イングランドのライム・リージス海岸や南ドイツを代表する多くの発掘地点で大量に発見される。
- 加えて、数メートルの地層の中で多様化も達成している。三畳紀の平らな二枚貝の場合は一つの種しか確認できないのに対し、ジュラ紀初期のアンモナイトには多数の種が存在し、その数も豊富であった。
- このことから、酸素濃度の大幅な低下が終息し、徐々に上昇に転じていたことが示される。

それにもかかわらず、酸素濃度がすぐに現在のようなレベルになったわけではない。アンモナイトが登場したのは、ジュラ紀初頭の海洋表層にわずかな酸素が溶け込み始めたからである。アンモナイトはこれを最大限に利用した。アンモナイトは、地球上で最も低酸素環境に適応した動物の一つと言えるだろう。そのため、ジュラ紀と白亜紀の温暖な海では生態学的に有利で、その利点を具体的に活用したのである。
ジュラ紀初期の地層から大量のアンモナイト化石が発見されることから考察すると、アンモナイトはその優れた構造により、海水に微量に溶けた酸素を最大限に抽出する能力を有していたと考えられる。言い換えれば、ジュラ紀から白亜紀にかけてのアンモナイトの体制は、三畳紀からジュラ紀の境界付近の全世界的な低酸素状態に対応すべく進化したものであった。また、鰓が非常に長いという特徴も有していた。
「アンモナイト」の鰓が『オウムガイ』(注167)のように四つだったか、現在のイカやタコのように二つだったかは定かでない。しかしながら、ジュラ紀初期のアンモナイトの大部分には流線形の殻が見られないことから、迅速に泳ぐ能力はあまりなかったと推測できる。迅速な泳動よりも、空気で充填された殻を飛行船のように利用し、海面近くでゆっくりと浮遊したり穏やかに泳いだりしていた可能性が高いのである。

(注167)軟体動物門頭足綱オウムガイ科に属する海産動物。「生きている化石」といわれる種の一つで、フィリピンを中心とした西太平洋の熱帯域に分布する。アンモナイトの生き残りないし子孫と誤解されることが多いが、そうではなく共通の祖先を持つ別種である。アンモナイト類とオウムガイ類は、どちらも房(小室)に分かれた殻をもっていて全体的に似ている。
ジュラ紀のアンモナイトは、白亜紀になるまで細部しか変化しなかったが、白亜紀に入ると殻の形状に目を見張るような変化が始まった。従来の平巻き型(殻が平面的に巻かれている)のもの(オウムガイと同様)が多く残されている一方で、他の形状の殻も現れるようになった。
アンモナイトは見た目上、オウムガイの殻にイカがはまり込んだような姿だったと推測される。現生のオウムガイには90本の触手があるが、アンモナイトの触手はおそらく8本か10本だったと思われる。また、オウムガイは腐肉を食べるのに対し、現在のイカや中生代のアンモナイトは肉食性で、食料として生きた生物を必要としていたである。

温室化した海を代表する第二の軟体動物は「二枚貝」である。形はアツバニマイガイほど奇抜ではないものの、現存するどの種類とも確かに異なる二枚貝、『イノセラムス』である。カキの近縁種であり、様々な種に多様化していたが、みな泥状の海底で競い合って生存していた。海底の表面に生息する特性から、どの種類も穴を掘る能力はなかった。
イノセラムスには極めて巨大なものも存在し、なだらかな肋(ろく、線状の高まり)の入ったそのアーモンド型の殻は、殻口から殻頂までの長さが2メートル以上にも達していた。しかしこの貝類は、現存するどの二枚貝とも異なり、大きさに対して殻が非常に薄い特性を持っていた。緩やかに隆起する上殻は、カキやイタヤガイ、コケムシなどの外肛動物、フジツボやチューブワームといった多様な動物に覆われることもあった。しかしほとんどのイノセラムスは、酸素が少なすぎて「普通の」軟体動物やその他の無脊椎動物が生存できないような海底や海中で生活していた。
他の二枚貝との大きさを比較するだけでも、イノセラムスがどれほど異なっていたかが理解できる。それらは重さ数百キロにも達する軟体部を支えていた。二番目に大きな二枚貝であるアメリカナミガイは、殻長がせいぜい30センチ程度で、軟体組織の重さは450~900グラム程度しかない。カキでも殻長30センチになるものはいるが、その数は多くない。イノセラムスは、巨大なオオシャコガイと比較的小さいアメリカナミガイの間の大きさに位置していた。
ペルム紀に登場してから白亜紀末に絶滅するまでの間に、数多くのイノセラムスの種類が存在した。その繁栄の舞台は、温室化した海であった。海底は低酸素で有機物に富み、そこからはメタンなどの化学物質が滲み出ていた。イノセラムスは、これらの化学物質を栄養に利用し、巨体を維持するために微生物を共生させていた。これは現生二枚貝が海水から食物を濾し取って食べる方法とは大きく異なる特性であった。
温室化した海についてもう一つ注目すべきは『中層水域』である。太陽光が届かない深さでありながら、淀んだ海底よりは何百メートルも上の海域である。現在、この広大な中層水域は地球上で最大の単一生息環境となっており、多様な生物の生息地となっている。海面で太陽光や大気に触れることも、海底に出くわすこともない環境に生物たちは適応している。こうした「中間」での生活が生命線となっている。
これらの生物にとっては、捕食の危険性や温度、酸素濃度の観点から、温かい浅海も冷たい深海も命取りとなる環境である。そのため、浮きも沈みもしない「中性浮力(浮きも沈みもしない状態)」を達成・維持できる適応構造を持つことが、生存のために最も重要である。例えば、現代の中層水域に生息する比較的大きな動物で最も一般的な「イカ」は、浮動のための触手を発達させている。また、体内に液胞を進化させ、その中に脂肪や塩化アンモニウム溶液などの化学物質を蓄え、それによって体全体の比重を海水より軽くしている。
イカの獲物は個々には小さいが、その量は膨大である。それは多種多様な浮遊・遊泳動物が密集したもので、その総称として『深海音波散乱層(DSL)』と呼ばれている。DSLは、1940年代に初めて使用されたソナーによる水深測定で発見された。DSLには小型の節足動物(甲殻類、端脚類(注168)、等脚類(注169)など)をはじめ、多様な動物が集まっている。「門」と呼ばれる分類群の動物が無数に存在する。日中は岸から遠く離れた海域で、この巨大な生物の層が水深約600~800メートルの位置に形成され、縦・横・深さの三次元で数百~数千キロもの大きさに広がっている。しかし、日が暮れるにつれて層全体が浅海に向けてゆっくりと上昇し始め、完全に暗くなると、何十億トンもの無数の動物がさらに浅く、温かく、栄養豊かな海域に達する。この深さには魚やイカなど視覚が発達した捕食者が存在するため、日中にここに来るのは命取りである。
(注168)体は一般に左右に扁平で、頭部、八胸節、七腹節から成り立っており、眼には柄がない。体長は0.5~2cmのものが一般的で、一部は淡水・陸上に生息するが、大部分は海生である。ハマトビムシ・ワレカラなど約5700種が存在している。
(注169)体長は約1cmのものが多く、体は背腹に扁平で、7胸節・6腹節・腹尾節から成り立っている。海生・淡水生・陸生のほか、寄生生活をするものも存在する。キクイムシ・シオムシ・ワラジムシなどがいる。
この新たな生活形態が白亜紀に登場したことは、豊富な証拠によって裏付けられている。それ以前の時代では、中層水域に追求すべき食物資源は存在しなかったと考えられている。その結果、生涯を浮遊しながら過ごし、夜ごと数百メートル上昇し、明け方に深層へと戻るという大規模な適応を遂げた種は存在していなかった。
しかし、中層水域にすむ節足動物の出現で急速な進化が進んだ結果、新たな仕組みの浮遊機能を使って浮き上がり、それらを捕食できる動物が誕生した。中層水域で無重力状態になる何らかの方法を身に付けることが、最も重要な適応だったわけである。
中層水域の食物資源を利用して進化した肉食動物は、主に『アンモナイト』であった。しかし、その形状はそれまでの先祖伝来のものとは大きく異なっていた。
- 海底近くに生息するアンモナイトが平巻き型である一方で、中層水域のアンモナイトは一生を浮遊して過ごすための特殊な殻を持っていた。これは奇妙な形状で、速く泳ぐことは不可能であった。
- しかしながら、一旦DSL(深層魚群)の生物が密集する水塊に入ると、食糧が豊富に存在するために、そこに留まることが可能であった。DSLが上昇すれば一緒に浮き上がり、日中は共にゆっくり沈んでいくだけで、食物に恵まれ、天敵のいない生活が可能であった。アンモナイトはこれにより平穏な浮遊生活を送っていた。大きな浮力装置を上部に配置し、その下に小さな乗員室を吊り下げる、まるで熱気球のような構造であった。

中層水域のアンモナイトは浮力を効果的に調整しなければならなかった。現存するオウムガイの浮揚システムは比較的原始的で、動作に時間がかかることが確認されている。しかし、アンモナイトの浮揚システムは、全体的に見て、はるかに優れていた可能性がある。その要となるのが、美しい縫合線を有する複雑な「隔壁」であった。浮上したいときには、隔壁で区切られた気房から迅速に水を排出し、沈みたいときにはそこに素早く水を戻してバラストのように浮力を下げる。白亜紀後期に登場したこの新しいタイプのアンモナイトは、「異常巻き」と俗に呼ばれる。デボン紀に出現してから白亜紀末に絶滅するまでの間、アンモナイト本来の渦巻き型から逸脱していたからである。これは、約6000万年にわたるアンモナイト類の生息期間の中で唯一の例であり、チクシュループ小惑星が衝突してアンモナイトを絶滅させるまで存続していた。

異常巻きアンモナイトの中には、巨大な巻貝に似たものも存在した。しかし、その殻はいくつもの気房に分割されており、最後の長い房は下向きに開き、触手などの軟体部を収納していた。また、巨大なクリップのような形状や、単純な大きな鉤状のものも存在した。しかしながら、
最も数が多かったのは長くて直線的な円錐形であった。円錐の尖っているほうが最初に形成される房で、成熟すると全長が約180センチに達することもあった。もちろん殻は浮揚のためにいくつもの気房に分かれており、そこから触手の先をまっすぐに垂らし、殻を垂直にして水中を浮遊していた。この種のアンモナイトは『バキュリテス』と呼ばれ、白亜紀後期の肉食動物としては地球上で最も個体数が多かった可能性がある。
中層水域はこれらバキュリテスの大群で溢れていた。白亜紀の海を描いた壁画や絵画にもよく登場するが、その姿は大抵間違えて描かれている。それらの描写では魚やイカのように、細長い体を水平に保つ生活様式が描かれているが、これは実情とは違う。実際には、最初に形成される小さな殻の部分を上にし、重い頭部や触手を下に垂らすように、体を垂直に保つ生活様式をとっていた。横向きに泳ぐことは不可能であり、体を横に保つことすらできなかった。すべてが上か下の方向を向いていたのである。動きは驚くほど速かったと思われ、水を噴射することで上方へと飛び出し、その後ゆっくりと下降していた。バキュリテスを捕食していた生物(おそらく魚やサメ)は、通常の攻撃を仕掛けると、そのたびにその動きに翻弄されたに違いない。通常なら、獲物は捕食者の前方へと逃げようと泳ぐ。しかし、バキュリテスに襲いかかると、操り人形のように素早く上方へと消えてしまう。それがバキュリテスにとっては前方であり、逃げる獲物がとるべき行動としては、全く正しいものであった。
中生代海洋大変革
中生代の後期に、海洋生物の攻防に画期的な変革が生じた。カリフォルニア大学デイヴィス校の古生物学者ヒーラット・ヴァーメイは、これを実に端的に『中生代海洋大変革』と呼んでいる。
- その大転換とは、海洋の捕食者が驚異的な進化を遂げたことに他ならない。ペルム紀以降の捕食者は、攻撃性を次第に増して、殻を破壊する能力を発達させていった。
- さらに、植物食性の無脊椎動物と比較的小型の肉食動物もそれに呼応して進化を遂げ、石灰質の外骨格をますます強化していった。これは現在、「中生代海洋大変革」として広く認識されている。
最初のうちこの表現は、ペルム紀末の大量絶滅後に捕食の手段が殻の破壊に向かったことだけを意味していた。古生代の二枚貝や巻貝、棘皮動物、腕足動物の殻はかつては難航不落の要塞であったが、殻の中にある栄養豊富な肉へと到達する新たな方法が進化したのである。しかしこの概念はその後拡張された。
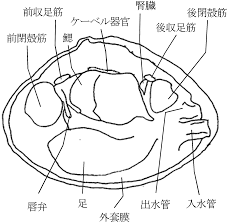
被食者側の適応も負けず劣らず見事である。以前は海底の表面やその少し下だけで暮らしていた二枚貝が、深い穴を掘る構造に進化した。この新しい二枚貝は、殻を噛み合わせるための鉸歯(こうし:二枚貝の殻と殻を合わせるための歯)に形の異なる主歯と側歯があるために、『異歯型』と呼ばれる。これらは大がかりな改造を行ない、「外套膜」(軟体動物の体表を覆う筋肉質の膜)の一部を癒合させて一対の水管をつくった。穴を掘るこのタイプは現在の二枚貝類の中でも最も多様化しており、どれも砂や泥やシルト(沈泥(ちんでい)、砂より小さく粘土より粗い砕屑物)の中に素早く潜り込むことが可能である。穴を掘る理由はただ一つ、捕食を免れるためであった。堆積物の上ではなく中にいれば、摂食の効率は必ずしも上がらないものの生存確率は大幅に向上する。二枚貝以外で、穴を掘る生活様式に適応すべく体形を大きく変えたものには、巻貝類、新種の多毛類(一般に細長く一様な多くの体節に分かれた動物で、ゴカイ類とも称される)、一部の魚類、まったく新しい種類のウニ類などが存在する。
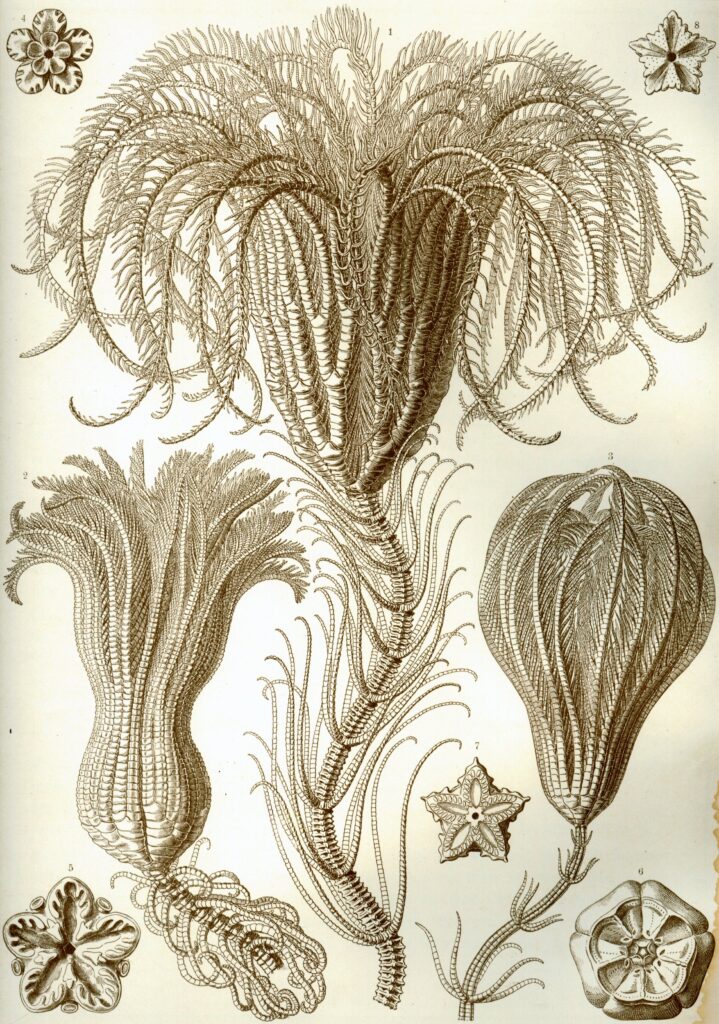
革新的な大転換を示した無脊椎動物は他にも存在する。『ウミユリ類』と呼ばれる棘皮動物の一群である。見た目は大きな花のようだが無脊椎動物であり、古生代の生物らしい固着性を持っていた。即ち、浮遊する幼生期を経て一度どこかに定住したら、生涯を通じて移動ができずに海底に固定される特性であった。現代のアメリカ中西部を車で走ると、かつてどれほど多くのウミユリが生息していたかがよく理解できる。どの崖の壁にも、ウミユリの長い茎部を構成する小さな丸い「骨片」が数えきれないほど詰まっている。これほどの量の化石が残っているということは、浅くて水の綺麗な暖かい海が広がっており、海底はウミユリで満ち溢れていたと推測できる。浅いとはいえ、海底まで太陽光が届いていたかどうかは不確かである。しかし、ウミユリにとってはそれでも問題なかったのであろう。獲物は微小なプランクトンであり、少なくとも代謝に関しては「スローライフ」を送っていた。ただし、一度固着した後は移動ができず、嵐や捕食者によって引き剥がされた場合、直ちに死ぬ可能性があった。
中生代海洋の大転換は、捕食者と被食者の変化だけでなく、新たな生息環境の大幅な拡大を意味している。これには、捕食から逃れるためにさらに深く穴を掘るように二枚貝や巻貝が形態を進化させたことや、堆積物を食物として利用する無脊椎動物が次第に増加したことも含まれている。このような変化が生じたことは、生痕化石の種類や数が増えていることから裏付けられている。これはカンブリア爆発の章で説明した状況と同様である。その結果、中生代の堆積物は生物によって徹底的に混乱させられることとなった。
中生代の海洋で動物に大きな転換が起こったのは、海底やその下だけではない。動物の誕生以来初めて、海面から海底までの全水域が大規模に利用されるようになった。
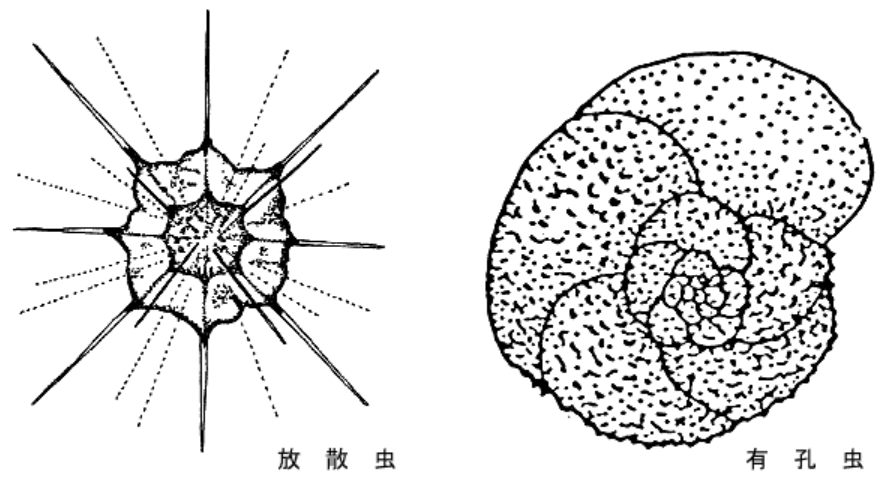
- 新たに現れた生物の多くは一般的な動物ではなく、主に原生動物や単細胞の植物プランクトンであった。中生代の地層からは、新しく出現した重要な生物群の微化石が発見されている。
- その一つが、多種多様な有孔虫類である。『有孔虫』はアメーバに似ているが殻をもち、海底に生息するもの(『底生有孔虫』)と、それよりはるかに上の水中に漂うもの(『浮遊性有孔虫』)もいる。
- 浮遊性生物(プランクトン)には、ほかに珪質の骨格をもつ『放散虫』という存在も確認できる。
しかし、中生代以後のプランクトンに起きた最大の変化は、『円石藻類』(<物語り その6>図66)と呼ばれる藻類の出現であったかもしれない。円石藻類の微小な骨格が海底に堆積して岩石化すると、我々がよく知る『チョーク(白亜)』になる。石藻はきわめて小さな植物であり、球体に近い体の表面に『円石』と称される円盤を6~12枚程度保有している。円石は炭酸カルシウムで成り立っており、その小ささゆえに顕微鏡なしでは観察することができない。円石藻が死ぬと、この微小な円石が海底に沈降し、無数に積み重なって、やがてかの有名な『ドーバーの白い崖』のような壮大な堆積岩を形成する。
ブリテン島からフランス、ポーランド、ベルギー、オランダ、スカンジナビア全域を経て、旧ソビエト連邦の大部分まで、そして最後に黒海に至るまで、ヨーロッパ北部の地層全体には、このような崖が連続して存在している。円石は地球の温度にも重大な影響を及ぼしてきた。その白さが太陽光を宇宙に反射し、結果として地球を冷却する効果がある。
カンブリア爆発において、動物は呼吸器系を中心に新たな体制を生み出した。そのように、三畳紀の海洋動物も新しい適応を多数示している。前章で説明した通り、陸上動物は様々な種類の肺を試した。同様に、海においても新たな試みが行われていた。例えば、軟体動物の二枚貝類は、新しい体制や生理機能の進化を遂げた。これは無限に広がる海底を利用するための適応であった。そして、その海底は栄養が豊富だが酸素濃度が低いという特徴を持っていた。
思考を深めると、酸素が不足していたからこそ、海底は二枚貝にとって最適な居住地となったとも言える。海底には、プランクトンなどの生物の死骸という形で、還元された炭素が大量に降り積もっていた。もし海底に酸素が存在していたなら、フィルター摂食生物や、堆積物や死骸を食べる生物によってこの物質はすぐに消費されたであろう。しかし、低酸素状態ではこうした生物が生息できず、普段は海底の死骸を分解している細菌でさえ寄りつけない状態であった。
こうした状況から生じた炭素循環の乱れが、三畳紀に酸素濃度が急落した一因である。しかしながら、二枚貝類はこの状況を乗り越える方法を見つけた。前述のイノセラムスのような、何らかの酸素が存在する海底に生息する種は、降り積もる有機物ではなく、有機物が豊富な堆積物の一部から滲み出すメタンを食物とした。メタン菌は低酸素、あるいは無酸素状態で繁殖する細菌である。わずかな酸素が存在する海底でも、堆積物に数センチほど潜ると無酸素地帯となる。このため、メタン菌が生存可能であった。メタン菌は代謝過程で副産物としてメタンを放出する。
二枚貝類は、メタンやその他の溶存有機物質を利用できる別の細菌を鰓に共生させていた可能性がある。もしくは、単にその細菌を餌としていた可能性もある。現在でもこれに似たメカニズムが、深海の熱水噴出孔周辺の動物相に見られる。巨大なチューブワームや二枚貝がこのような化学物質を栄養源としている。ただし、現代の熱水噴出孔と中生代の二枚貝類との間には、酸素濃度が高いという違いがある。熱水噴出孔で生活する生物には鰓が必要ない一方、中生代の二枚貝類はそうはいかなかった。
さらに、危機的な酸素不足に適応して、二枚貝類とは異なる形態を進化させた生物も存在する。それが『カニ類』と『ロブスター類』である。全体が小エビのような形の甲殻類は古生代の岩石からも発見されるが、カニの形状は比較的新しい進化である。カニは形状的にはエビに似ているが、ただ腹部が胸の下に折り畳まれているだけである。頭部と胸部は融合し、石灰質で強固な甲羅を形成しているため、天敵から見て手強い存在である。腹部を甲羅の下に収めるという設計は非常に優れている。どのような攻撃を受けるにせよ、腹部が一番狙われやすい。そのため、腹部を甲羅の下に保護することでこの弱点を排除し、カニは海洋生態系で急速に影響力を増した。
さらに、大きなハサミを持つことで硬い軟体動物の殻を割り開くことが可能になった。それまでは、殻を持つ生物の中身を食べられる捕食者はほとんど存在しなかった。しかし、カニを含む甲殻類はその手段を見つけたのである。
カニの形態は非常に革新的ではあるが、その形態を取る背後に特別な理由があるわけではない。一般的には防御(腹部を折り畳み、頭胸部を石灰化して分厚い甲羅を形成する)と攻撃(強力なハサミの進化)の観点から説明されている。しかしながら、別の見方も存在する。カニの形態の出現は、呼吸の効率を向上させるためだという説がある。頭胸部の下に空間を確保し、鰓を囲み、その鰓に水を供給するポンプを発展させることで、呼吸効率が向上したのである。
カニはエビに似た生物から進化した存在である。これらの祖先の形態を遡ることにより、カニの鰓の構造へ至る進化の過程が明らかになる。エビでは、鰓は体の下部に位置しているものの、全体が完全に覆われているわけではない。背側は保護されているが、鰓は体節に接しており、下側は水に直接触れている状態である。
中生代の海洋は温暖化の影響で時間とともに変化していった。それにもかかわらず、当時を象徴する「アンモナイト類」と「イノセラムス類」は、状況が許せば今でも生存していた可能性がある。約6500万年前、チクシュルーブ小惑星の衝突が起きたその日が、中生代の独特な生態を持つ生物の絶滅を招く災厄であった。
第11章 恐竜の死【6500万年前】
恐竜が隕石衝突により滅ぼされたという「事実」が広く受け入れられるようになったのは1990年頃からであった。これはカリフォルニア大学バークレー校のアルヴァレスのグループが衝撃的な発見(注170)をした後の10年間で、この発見は大量絶滅だけでなく地質学的なプロセス全般に対する我々の理解を根本から変えるものであった。
(注170)L. W. Alvarez et. al., “Extraterrestrial Cause for the Cretaceous-Tertiary Extinction”, Science 208, no.4448, 1095 (1980)
新世代の地質学者や博物学者の間では、「斉一説」(注171)と「天変地異説」が交互に優位に立つことがあった。斉一説が最終的に優勢になったのは、岩石の特性や年代を解析する技術が進歩していた一方で、地球規模の大異変の証拠が一度も見つからなかったからである。それどころか、繰り返し大異変に襲われた形跡も見つからなかった。しかし、大異変が反復して起きなければ説明がつかない事象も存在した。なぜなら、すでに複数回の大量絶滅が過去に発生したことが確認されていたからである。
(注171)自然において、過去に作用した過程は現在観察されている過程と同じだろう、と想定する考え方。「現在は過去を解く鍵」という表現で知られる近代地質学の基礎となった地球観。天変地異説に対立する説として登場した。
その大量絶滅とは、古いものから順に、『オルドビス紀』、『デボン紀』、『ペルム紀』、『三畳紀』、『白亜紀-第三紀境界』で起きたものであり、現在これらは『ビッグファイブ』と呼ばれている。
20世紀の初頭には、天変地異説を支持する者はもはや存在しなかった。斉一説を唱える者たち(チャールズ・ダーウィンを含む)は勝利を確認したものの、大量絶滅の説明については依然として疑問を抱えていた。
地質学者たちは、大量絶滅が非常にゆっくりと確実に進行したという結論を下し、観測可能な気候変動や海水面の変化も、適切な時間を経れば種の絶滅を引き起こす要因になり得るとした。ビッグファイブは、20世紀後半にはすべての地質学者に受け入れられていた。1950年からの30年間、地質学は停滞を続け、すべての現象が地球上で長期間進行する過程によって説明可能だという認識に自己満足していた。しかし、その状況は1980年6月6日に一変した。
1980年6月6日、ノルマンディ上陸作戦36周年の日に、アルヴァレスが「『K-T境界の大量絶滅』は『隕石の衝突』によって引き起こされた」と主張する論文が公表されたのである。
当時の斉一説は、長い歴史と威厳を持ちながらもすでに揺らぎ始めていた。アルヴァレスの論文は、その斉一説という巨大な体系全般への、とくに大量絶滅の原因についての定説に対する攻撃にほかならなかった。
隕石衝突と大量絶滅
固体表面を有する太陽系内のすべての惑星と衛星には、数多くの衝突クレーターが存在している。これは太陽系の初期に隕石衝突が頻発し、その影響が非常に大きかったことを示す証拠である。隕石衝突は、太陽系外でも全ての惑星系にとって危険な存在であるといえる。惑星規模の大変動の中でも、隕石衝突は重大な出来事として頻繁に発生している。隕石衝突によって、優勢だった生物群が排除されると、新種の生物や以前は目立たなかった種が台頭し、惑星の生物史が劇的に変化する可能性がある。このような観点から、1980年のアルヴァレスの論文は特別な意義を持つものとなった。
「K-T境界絶滅」が大型天体の衝突によって引き起こされたと考えられるようになったのは、以下の二つの重要な発見が証拠として示されたからである。
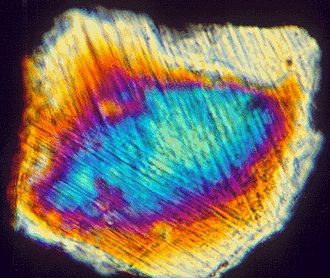
- 一つは、境界粘土層で『イリジウム』値が増加していること
- もう一つは、イリジウムに交じって大量の『衝撃石英』が見つかったこと
1997年時点で、全世界で50箇所以上のK-T境界層で高濃度のイリジウムが検出されていた。イリジウムは地表にはごくわずかしか存在せず、小惑星や彗星には地球よりもはるかに高い濃度で存在することが多い。
- そのため、高濃度のイリジウムは隕石衝突の証拠と見なされている。
である。
また、「衝撃石英」の粒子が隕石衝突の指標とされる理由は、多くのK-T境界層で特徴的な縞模様を持つ石英が観察され、
- その形状が非常に高い圧力を受けた場合にのみ形成されるためである。たとえば、石英を含む岩石に、大型の小惑星が高速でぶつかるといった場合である。
地球の条件下では、石英粒子にそうした縞模様が自然に生成されることはない。
さらに、K-T境界層には、「衝突直後に起きたと思しき大火災の痕跡」も残されている。世界各地のK-T境界粘土層からは「煤」の微粒子が発見されており、この種の煤は植物が燃えるときにのみ生成される。
- この煤の量から推測すると、地表を覆っていた森林や低木地帯の大部分が火災により焼き尽くされたと考えられる。
アルヴァレスの仮説は初めのうちこそ議論を呼んだものの、1980年代を通じて鉱物学、化学および古生物学の分野から裏づけとなるデータが次々に集まった。
その結果、
- 大型(最大で直径10~15キロメートル)の彗星か小惑星が6500万年前に地球に衝突し、当時地球上に存在した種の半分以上がK-T境界期に急激に絶滅したという事実をほとんどの専門家が認めるようになった。
- メキシコのユカタン半島で発見された大型の『衝突クレーター(チクシュルーブ・クレーター)』はこの時代に形成されたもので、まだくすぶっていた衝突仮説への反論を大幅に減少させた。
アルヴァレスらは、『ブラックアウト』と呼ばれる長期間にわたる真っ暗な状態が生物の絶滅を決定的にしたと主張している。大衝突の後、隕石と地球の物質が大量に飛び散ってブラックアウトが起き、それが長期に及んだことで、プランクトンを含む地球上の植物の多くが根絶やしにされた。植物が死滅したために、災いと飢餓の波は食物連鎖の階段を上へ上へと広がっていった。
いくつかの研究グループがモデルを作製し、そうした大気変化が生じた場合にどのような致死的状況が訪れるかを割り出した。その結果、
- 大量の硫黄が大気中に放出され、一部が硫酸となり、『酸性雨』として地表に降り注いだと考えられる。酸性雨が直接生物に影響を及ぼした可能性もあるが、より大きな影響はその『冷却効果』であった。
- 生物圏にとってさらに深刻だったのは、『地表に届く太陽エネルギーが減少』(8~13年間で20パーセントもの減少)であり、これは、『大気中の塵粒子(エアロゾル)』によって太陽エネルギーが吸収されてしまった結果である。
- これにより、衝突時にはおおむね熱帯性気候であった世界が氷点下かそれに近い低温にまで冷え、それが10年間続いたとしてもおかしくない。
- このモデルの結論は、ブラックアウトが大量絶滅の一因だとするアルヴァレスの当初の主張、つまり大気中のエアロゾルの量が急激に増えることが長い冬を引き起こしたという仮説を裏付けている。
衝突後には大量の塵が発生した。大型(直径10キロメートル)の小惑星や彗星が衝突すると大気中に大量の塵が放出され、地球の気候に様々な影響を及ぼす。その一つが長期間(数か月程度)のブラックアウトであり、これにより光の量が不足して光合成に必要なレベルを下回り、さらには陸地が急速に冷えた。
しかし、
- このモデルが導き出した最も不吉な予測は、その塵が『水循環にも変化』をもたらしたことである。これは、以前に注目されていなかった側面である。モデルによれば、地球全体の平均降水量は数か月にわたって九割以上減少し、衝突後一年が経過した時点でもまだ正常時の半分程度にしか戻らなかった。
言い換えるなら、地球は『寒くなり、暗くなり、そして乾燥した』のである。これらが揃った結果、大量絶滅の条件が整ったとも言える。特に植物と、植物を食べる動物にとっては。
モデルが示したのはこれだけではない。
- 衝突から数時間のうちに、大気圏外ぎりぎりの高さにまで岩石の破片が吹き飛ばされ、それが再び高速で落ちてきて地上に降り注いだ。岩石は非常に高温になっていたため、地上の植物が燃え始めた。結果として、「史上最大の森林火災が全体陸で発生した」のである。
これだけの事象が発生した結果、陸上の恐竜が全滅したという事実は全く驚くべきことではない。
先行する絶滅
K-T境界絶滅では、「全生物種の75パーセントもが死滅した」という事実が明らかになっている。
- 陸上では恐竜が消滅する一方で、哺乳類が台頭した。
- 海では、境界の白亜紀側でアンモナイトが姿を消し、古第三紀側では新たな海洋動物群が現われ、二枚貝や巻貝がその多数を占めた。
当初の仮説では、K-T境界で一度きりの『大量絶滅』が起きたとされていたが、年代測定の方法が進歩するにつれ、実際はそれよりはるかに複雑だったことが明らかになりつつある。
- 近年では、最後の一撃の前に少なくとも二回の『前K-T絶滅』があったことが確認されており、これに対する異論は存在しない。
近年の研究から、
- 生物が死に至る過程で火山活動による『洪水玄武岩』(注172)の影響もあったと再認識されている。
(注172)地中から比較的短い期間で非常に膨大な量の玄武岩質熔岩が噴出し形成される玄武岩の巨大な岩体のことである。その地形から「台地玄武岩」とも呼ばれている。カンブリア紀以後でも何回かの洪水玄武岩の形成が発生している。
ところで、恐竜の化石は数が非常に少ないため、化石を利用して恐竜が滅びていった速度を調べることはほぼ不可能である。その代わりに、「微化石」は豊富に見つかっており、隕石衝突による急激な絶滅説の裏づけはこの微化石の研究から得られた部分が大きい。しかし、依然として陸でも海でも大型動物がどうなったのかを解明する必要性がある。そのために最も詳しく研究された海の化石が頭足類のアンモナイトである。そして、このアンモナイトの調査の過程で、アンモナイト類の急激な絶滅の200万年前にイノセラムス類の二枚貝が徐々に死滅していったことが明らかになった。この二枚貝の死滅には興味ある事象が伴っていたので、ここではそれを紹介する。
ビスケー湾に沿った厚い地層の「露頭」(注173)は、白亜紀後期の赤道付近に生息していた最後のアンモナイトの絶滅状況を調べるのには絶好の場所である。この露頭は、フランス南西部からスペイン北東部までの広大な領域にまたがっている。特に、バスク地方のスマイアという古い村の近くにある岩の切り立った海岸線は、7200万年前から5000万年前までの地層が数百メートルにわたって続いており、本のページを開いたような形状を呈している。大量絶滅が起きたK-T境界の地層は岩に囲まれた入り江に位置し、岩石の種類と色も変化しているため見逃すことはない。
(注173)岩石や地層が土壌や植生などに覆われないで、直接地表に現れているところ。海岸や河岸などには露頭がよく発達するほか、採石場や道路沿いの崖にも人工的な露頭がみられる
ビスケー湾の研究と、他の白亜紀後期の地層での調査を踏まえると、アンモナイト類の急激な絶滅の200万年前にイノセラムス類の二枚貝が徐々に死滅していったことが明らかになる。カリフォルニア大学バークレー校のチャールズ・マーシャルとピーター・ウォードの研究では、マーシャルが開発した統計的手法を用いて調査した結果、隕石衝突の最も重要な証拠を含む地層に至るまで、アンモナイト類の少なくとも22種が存在したことが明らかになった。その証拠とは、イリジウムと衝撃石英、そしてガラス質の小球(注174)である。
(注174)「テクタイト」と呼ばれ、隕石落下の衝撃で上空へ吹き飛ばされた岩石の破片が、微細なガラス質となって再び高速で地上に降り注いだものである。
イノセラムス類の絶滅が興味深いところは、アンモナイト類よりかなり早い時期に消失したという事実ではない。注目すべきはその絶滅が多種多様な地域と時代において発生したという点である。例を挙げると、
南極の白亜紀の岩石に含まれる最後のイノセラムス類は約7200万年前(アンモナイト絶滅のおよそ700万年前)のものであった。イノセラムス類は広範に分布していたにもかかわらず、結局は全てが消滅の波に飲み込まれていったことが現在確認されている。
- この波は南極地域から始まり、その後徐々に北半球に移動した。病症が北方向へと進行し、徐々に二枚貝を死滅させるかのように。しかしこの場合、病症ではなく、「寒さ」と「酸素」が原因であった。
- 白亜紀の終わりに近い南半球の高緯度地域で、『熱塩循環の酸素版』ともいうべき現象が発生し始めた。通常の熱塩循環には温度と塩分濃度がかかわるが、この場合は温度と酸素濃度が重要であった。
- 高緯度地域で冷却された高酸素の海水が深層に沈むと、それが約200万年をかけて南から北へと移動し、全ての海洋に広がった。この水塊の存在が、イノセラムス類の二枚貝の終焉をもたらした。
- この二枚貝の消失は、生命史の中でも注目すべき出来事である。なぜなら、その時点までは1億6000万年以上ものあいだ繁栄していたからである。しかし、イノセラムス類が適応していたのはこの種の海ではなく、「低酸素で温かいタイプの底層水」だった。だから「寒さと酸素」に息の根を止められたのである。
以上が、生物史の中でも顕著な出来事であるイノセラムス類の二枚貝の消失に関する議論であった。次に、「K-T境界絶滅」へと戻ることにしよう。K-T境界絶滅を引き起こしたと推定される大元の事象について、現時点で解明されている事項を整理する。
- 隕石が衝突したのは一回だけであり、それは世界規模で海水面の急激な変化が二度発生したすぐあと(100万~300万年後)のことであった。
- その二度の変化に挟まれるようにして、海水の水質も大きく変化した。
- 衝突によって生まれた大きな(最大直径は300キロ)クレーターは、現在は『チクシュルーブ』と名づけられ、ユカタン半島に位置している。
クレーターの正確な大きさについてはまだ議論があるものの、その構造がクレーターであるということについては異論がない。絶滅の規模があれほど大きかった理由としては、衝突地点の地質と地形が関与した可能性がある。
というのも、衝突地点には硫黄含有量の多い蒸発岩(水溶液から水分が蒸発し、溶解していた物質が析出沈殿してできた堆積岩)が存在するうえ、飛来した隕石自体の内部にも硫黄が含まれていたことが、衝突後の大気の致死性を高めたおそれがあるからである。
- 6500万年前の隕石は、赤道付近の浅い海に衝突した。その海底には、硫黄を豊富に含む蒸発岩が堆積した炭酸塩プラットフォーム(炭酸塩が堆積した平坦な地形)があり、結果的に信じ難いほど悲惨な状況が生じたと考えられている。
- 例えば、世界規模での大気成分の変化や、それに伴う気温の低下、酸性雨(おもに衝突地点に存在した蒸発岩由来の硫黄による)、世界各地で発生した大火災などが、絶滅につながるメカニズムとして考えられている。
メキシコの東海岸には、ごつごつした砕屑性の厚い堆積岩層がいくつも存在するが、これらが衝突の衝撃波によって形成されたとする見解が大多数の科学者たちによって共有されている。
このように、隕石の衝突が引き起こした長い冬が、絶滅へと至る最も大きな影響を及ぼしたとされている。この冬は、大気中のエアロゾルが短期間で大幅に増加した結果、引き起こされたと考えられている。
近年、衝突後の大気の変化を模したモデルが発表された。それによると、衝突によって大気中の塵の濃度が著しく上昇したことも、生物にとって致命的だった可能性がある。
- 微細な塵が長期(数か月程度)にわたるブラックアウトを生み出した結果、光の量が減る(光合成に必要なレベルを下回る)とともに、陸地の気温が急降下した。こうした大量の塵は地球の水循環にも深刻な影響を及ぼした。
- 高度な気候モデルによれば、大型隕石の衝突後には世界の平均降水量が数か月にわたり90%以上減少し、衝突後1年経過しても正常時の半分程度しか回復しなかった。それが生物相に影響することは、現在では十分に立証されている(注175)。
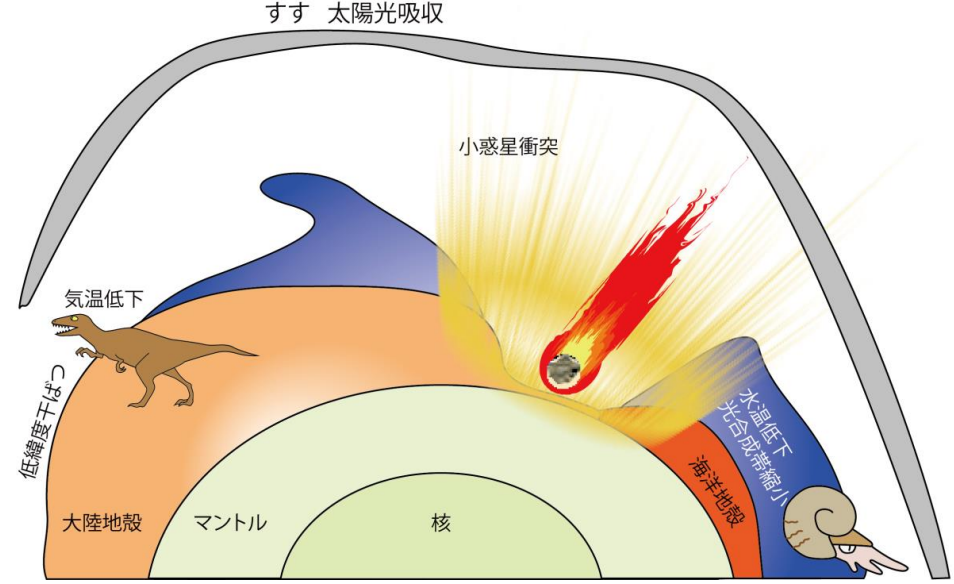
(注175)【恐竜やアンモナイト等の大量絶滅のストーリー】(東北大学大学院理学研究科・気象庁気象研究所)の説:小惑星衝突が有機物を燃やし、すすを成層圏に放出、太陽光吸収により、気温低下、低緯度での干ばつ、海水温度の低下、海洋光合成帯の縮小を起し、それにより恐竜やアンモナイトなどが絶滅した。
デカントラップの洪水玄武岩による温室効果絶滅
しかし、地球に小惑星が衝突した際には、すでに『洪水玄武岩』が火山活動によって噴出している最中であった。これは地球史上における稀有な大規模な出来事であったことが明らかにされている。この事件の結果、莫大な量の玄武岩が地球の深部から噴出した。この時期の洪水玄武岩が形成した高原は『デカン・トラップ』と称される(注176)。
(注176)Blair Schoeneらはウラン・鉛(U-Pb)年代測定法を用いてデカン・トラップでの噴火を正確に時系列でまとめた(注177)。U-Pb年代測定法では、通常は冷えていくマグマの中で形成される鉱物であるジルコンの個々の結晶内のウラン鉛同位体比を測定することで、以前に使用されていた技術よりはるかに正確な年代を割り出すことができる。新たにネジを巻いた時計が正確に秒を刻むように、ジルコンに含有されたウラン放射性同位体は、ジルコンが結晶化するとすぐに崩壊しはじめ、ゆっくりと確実に鉛同位体へと変化する。その速度は分かっている。ゆえに、個々のジルコン内の2つの同位体の現在の比率を使って、誤差±40,000年という精度で年代が測定できる。Schoeneらはデカン・トラップで最も大きく完璧な地層の9つを試料とし、古代の溶岩流の間に挟まった火山灰層と土壌からジルコンを収集した。この方法によって各噴火イベントの年代を決定することができ、その結果、
- デカン・トラップでは4回の大規模噴火があり、それぞれが約100,000年間続き、大量のマグマと気候変動を引き起こす温室効果ガスが周囲に放出されたことが示された。
- この結果は、これらの噴火イベントがチクシュルーブ小惑星衝突の数万年前に始まったことを示している。
- Schoeneらは、デカン高原での噴火で大気中に放出されたメタン、二酸化炭素、二酸化硫黄のせいで地球の気候と環境が激変し、大量絶滅が誘発され、それはチクシュルーブ小惑星衝突のかなり前だったとしている。
(注177)Blair Schoene et al., SCIENCE 363 (2019) 862-866.
おそらく8400万年前には、広範な溶岩がコア・マントル境界近くから上方へと移動し始め、約2000万年をかけて地表に至ったと考えられている。これほどの大量の溶岩が動くと、『真の極移動』(<物語り その8>図91)を引き起こす可能性が高い。真の極移動は質量分布の偏りが存在するときに起きる。それにより、自転する地球の慣性モーメントが変化し、大陸を移動させる。この急速な移動により、環境が不安定化した可能性がある。たとえば、カナダ西部とアラスカの大部分は、8400万年前まではメキシコと同じ緯度に存在したにもかかわらず、中生代末にはメキシコからはるかに離れていた。
- 洪水玄武岩の噴出は様々な影響を及ぼすが、特に生命にとって最も重大なのは、それに伴う二酸化炭素などの『温室効果ガス』の大量放出である。

実際にそのような事象が何度も起きていることが観察されている。南北の極地方や高緯度地域が急速に温暖化する一方で、赤道地方の変化はそれほど急激ではない。このような環境変化が『温室効果絶滅』を引き起こす。大規模な洪水玄武岩の噴出は高緯度地域を加熱し、海を静止させて海中の酸欠状態を引き起こす。有毒な硫化水素を多量に含む深海水が海面へ上昇し、生物はデボン紀、ペルム紀、三畳紀後期のように大量絶滅に遭遇する。
実際に、「デカントラップ」も大きな影響を及ぼしたことが明らかにされている。ピーター・ウォードらが南極で行った研究によると、隕石衝突の数十万年前に海洋の温暖化が確かに起こり、それによって絶滅した生物種も存在した(注178)。
(注178)白亜紀中頃はマントル・プルームの活動が活発になり、中央海嶺での海洋プレート生成が増大し、大量の二酸化炭素が放出された。この結果、強い温室効果が引き起こされ、地球全体が温暖化した。氷河が全て融け、無氷期が訪れた。海面水位は現在よりも約200メートル高く、平均気温は現在より10~15℃高かったとされている。
地球温暖化の程度は、洪水玄武岩が主な要因とされる中、特に高緯度地方で顕著である。熱帯地方はすでにその限界に近い温度に達しており、これ以上の暖化は困難である。現代の世界を観察すれば明らかなように、温度変化が最も直接的に影響を及ぼすのは北極と南極である。これらの地域では、温度の変動による大災害や生物の絶滅が起こりやすい。
K-T境界絶滅についても同様の状況が見られる。大型の小惑星が地球に衝突したのは確かであるが、それより数十万年前に洪水玄武岩による急激な温暖化が生じ、海が淀んでいたのである。
本章の締め括りには、使い古されたボクシングの喩えがふさわしい。最終的に相手をノックアウトするのは当然ながら一発のパンチである。しかし、試合開始直後の最初の一撃で相手が倒れることはめったにない。それがどんなに強力なパンチであっても。ジャブやボディブローを何ラウンドも打ち続けることで、ノックアウトの舞台が整う。同じように、
- デカントラップが世界を弱らせ、隕石がとどめを刺したのである。
図表
図198 パンゲア超大陸とその分裂
コトバンク「パンゲア」・デジタル大辞泉「パンゲア」の意味・読み・例文・類語(https://kotobank.jp/word/%E3%83%91%E3%83%B3%E3%82%B2%E3%82%A2-606287)
図199 裸子植物(左)と被子植物(右)の胚珠
Qikeru(キケル)
(https://media.qikeru.me/angiosperms-gymnosperms/)
図200 恐竜と鳥類の系統樹
川崎悟司@satoshikawasaki
(https://twitter.com/satoshikawasaki/status/1421722216253706245)
図201 トロオドンの想像図
丹波地域恐竜化石フィールドミュージアム「トロオドン類」
(https://tamba-fieldmuseum.com/study/fossil/troodon)
図202 ドロマエオサウルスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「ドロマエオサウルス」(https://ja.wikipedia.org/wiki/%E3%83%89%E3%83%AD%E3%83%9E%E3%82%A8%E3%82%AA%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9#:~:text=%E3%83%89%E3%83%AD%E3%83%9E%E3%82%A8%E3%82%AA%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9%EF%BC%88Dromaeosaurus%EF%BC%89%E3%81%AF,%E3%83%88%E3%82%AB%E3%82%B2%E3%80%8D%E3%81%AE%E6%84%8F%E5%91%B3%E3%81%A7%E3%81%82%E3%82%8B%E3%80%82)
図203 始祖鳥の想像図
フリー百科事典『ウィキペディア(Wikipedia)』「始祖鳥」(https://ja.wikipedia.org/wiki/%E5%A7%8B%E7%A5%96%E9%B3%A5)
図204 プテロダクティルスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「プテロダクティルス」(https://ja.wikipedia.org/wiki/%E3%83%97%E3%83%86%E3%83%AD%E3%83%80%E3%82%AF%E3%83%86%E3%82%A3%E3%83%AB%E3%82%B9)
図205 エオアルラヴィスの想像図
Wikipedia, the free encyclopedia“Eoalulavis”
(https://en.wikipedia.org/wiki/Eoalulavis)
図206 有羊膜類の卵の構造
生物学 第2版 — 第29章 脊椎動物 —,OpenStax, Rice University,6100 Main St., Houston,
TX 77005-1827にて公開されている教科書“ Biology 2e”の日本語訳(https://medium.com/@BetterLateThanNever/%E7%94%9F%E7%89%A9%E5%AD%A6-%E7%AC%AC2%E7%89%88-%E7%AC%AC29%E7%AB%A0-%E8%84%8A%E6%A4%8E%E5%8B%95%E7%89%A9-bb7412be3b72)
図207 ケツァルコアトルス(翼竜)
恐竜図鑑「ケツァルコアトルス」
(https://kyouryu.info/quetzalcoatlus.php)
図208 ゲルマノダクティルス(翼手竜)
フリー百科事典『ウィキペディア(Wikipedia)』「翼指竜亜目」(https://ja.wikipedia.org/wiki/%E7%BF%BC%E6%8C%87%E7%AB%9C%E4%BA%9C%E7%9B%AE)
図209 サンゴ全体とポリプの構造
日本サンゴ礁学会・サンゴ礁Q&A
(http://www.jcrs.jp/wp/?page_id=622)
サンシャイン水族館・いきものAZコラム企画『いきものがたり』サンシャイン水族館(https://ikimall.ikimonopal.jp/blog/%E3%81%84%E3%81%8D%E3%82%82%E3%81%AEaz%E3%80%80%E3%82%B3%E3%83%A9%E3%83%A0%E4%BC%81%E7%94%BB%E3%80%8E%E3%81%84%E3%81%8D%E3%82%82%E3%81%AE%E3%81%8C%E3%81%9F%E3%82%8A%E3%80%8F%E3%82%B5%E3%83%B3%E3%82%B7/)
図210 ラディオリテス(厚歯二枚貝)の想像図
古世界の住人
(https://paleontology.sakura.ne.jp/radhioritesu.html)
図211 モササウルス・ベアウゲイの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「モササウルス」(https://ja.wikipedia.org/wiki/%E3%83%A2%E3%82%B5%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9#:~:text=%E3%83%A2%E3%82%B5%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9%EF%BC%88%E5%AD%A6%E5%90%8D%3A%20Mosasaurus%EF%BC%89%E3%81%AF,%E3%81%97%E3%81%9F%E5%8F%AF%E8%83%BD%E6%80%A7%E3%82%82%E3%81%82%E3%82%8B%E3%80%82)
図212 ハロビアの化石
レッドデータブック2015「ハロビア」
(https://www.pref.kyoto.jp/kankyo/rdb2002/geo/db/soi0024.html)
図213 モノチスの化石
フリー百科事典『ウィキペディア(Wikipedia)』「モノチス」(https://ja.wikipedia.org/wiki/%E3%83%A2%E3%83%8E%E3%83%81%E3%82%B9)
図214 アンモナイト[左]隔壁と連室の切断面[右]中層の縫合線
フリー百科事典『ウィキペディア(Wikipedia)』「アンモナイト」(https://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%B3%E3%83%A2%E3%83%8A%E3%82%A4%E3%83%88)
図215 オウムガイ[左]と殻の切断[右]
フリー百科事典『ウィキペディア(Wikipedia)』「オウムガイ」(https://ja.wikipedia.org/wiki/%E3%82%AA%E3%82%A6%E3%83%A0%E3%82%AC%E3%82%A4#:~:text=%E3%82%AA%E3%82%A6%E3%83%A0%E3%82%AC%E3%82%A4%E3%81%AF%E3%82%AA%E3%82%A6%E3%83%A0%E3%82%AC%E3%82%A4%E7%9B%AE%20%2D%20%E3%82%AA%E3%82%A6%E3%83%A0%E3%82%AC%E3%82%A4,%E3%82%92%E6%8C%81%E3%81%A4%E5%88%A5%E7%A8%AE%E3%81%A7%E3%81%82%E3%82%8B%E3%80%82)
図216 イノセラムスの化石
フリー百科事典『ウィキペディア(Wikipedia)』「イノセラムス」(https://ja.wikipedia.org/wiki/%E3%82%A4%E3%83%8E%E3%82%BB%E3%83%A9%E3%83%A0%E3%82%B9)
図217 アンモナイトの隔壁
古世界の住人
(https://ameblo.jp/oldworld/entry-10639309091.html)
図218 異常巻きアンモナイト
三笠市立博物館・博物館ニュース・[プレスリリース]北海道羽幌町より新種の異常巻き
アンモナイトを発見
(https://www.city.mikasa.hokkaido.jp/museum/detail/00010197.html)
図219 バキュリテスの想像図
広島空港ソラミィパーク、ジェラシックエアポート、生命の歴史(https://www.hij.airport.jp/special/dinosaur2020/cambrian/cambrian08.html)
図220 二枚貝の体の構造
群馬大学教育実践研究 第31号 27~32頁 2014「中学校理科における軟体動物の体の特徴
の理解 1.二枚貝」
(https://gair.media.gunma-u.ac.jp/dspace/bitstream/10087/8389/1/NO31_2014_04.pdf)
図221 ウミユリの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「ウミユリ」(https://ja.wikipedia.org/wiki/%E3%82%A6%E3%83%9F%E3%83%A6%E3%83%AA)
図222 有孔虫と放散虫
神戸市教育委員会、デジタル化・神戸の自然シリーズ、神戸の自然シリーズ11
神戸港のプランクトン
(http://www2.kobe-c.ed.jp/shizen/plankton/plankton/11084.html)
図223 衝撃石英の偏光顕微鏡写真
フリー百科事典『ウィキペディア(Wikipedia)』「衝撃石英」(https://ja.wikipedia.org/wiki/%E8%A1%9D%E6%92%83%E7%9F%B3%E8%8B%B1)
図224 恐竜やアンモナイト等の大量絶滅のストーリー
東北大学・2016年 | プレスリリース・研究成果「恐竜やアンモナイト等の絶滅は「小惑星衝突
により発生したすすによる気候変動」が原因だった」
(http://www.tohoku.ac.jp/japanese/2016/07/press20160714-01.html)
図225 地球内部の溶岩の流れ
asahi.comトップ > 教育 > NIE > ののちゃんのDO科学記事(https://www.asahi.com/edu/nie/tamate/kiji/TKY200509260253.html)
