
第四部 生命の進化
第9章 低酸素世界における恐竜の覇権【2億3000万 ~ 1億8000万年前】
大量絶滅が発生した後の時代は、ジュラ紀も含めて一般的に、生物が短期間で急速に進化するパターンが観察される。これらの時代は回復期と呼ばれている。各回復期の始まりは、大量絶滅を生き延びた生物群のみでその多様性は低いが、わずか500万年から1000万年で回復期は終わる。そして絶滅の影響が落ち着くと、多様性は必ず再び増大に転じる。
新たに登場する動植物は、必ず多種多様な種から成り立っている。これらの種は大半が回復期に新たに誕生したものであるが、絶滅前の時代から生き残った種も存在する。これらの種はかつて個体数が少なく、不安定な生活を強いられていたが、絶滅後の新しい世界で爆発的に増加し、生態系で固定的な地位を獲得した。ジュラ紀の初期も例外ではなく、回復の過程で新たな海洋生物が続々と生まれた。その大部分は新種の軟体動物や海生爬虫類で、新種の硬骨魚も多数出現した。しかし、ジュラ紀も白亜紀も、海洋生物が主役ではなく、無論のこととして主役は『恐竜』であった。
恐竜
生命の歴史を語る以上、『恐竜』の記述は欠かすわけにはいかない。イギリスのヴィクトリア朝以来、人々の大きな興味を引き続け、すでに語り尽くされている感がある。しかし、21世紀に入ってからは、「恐竜」について様々な新発見が報告されている。
恐竜について概説するとき、通常、次の三つの点が中心となる。
- まず、「恐竜は恒温性であったのか、それとも変温性であったのか」。
- 次に、「どのように繁殖し、営巣行動については何が明らかになっているのか」。
- そして三つ目は、「結局、どのようにして絶滅したのか」、である。
しかし、注目すべき謎はこれだけではない。特に興味深いのは、
- 「そもそもなぜ恐竜が存在したのか」、という問いである。少なくとも、「ああいう体のつくり(体制)がどうして生まれたのか」は明らかにしたい問いである。
- この問いは、「どのように呼吸していたのか」という問いに関連している。本章で探究するもう一つの疑問もまた呼吸に関連している。すなわち、「恐竜から鳥への進化の過程で何が新しく明らかになったのか」である。
事実、相当な数の新たな発見があったが、その大部分は中国の調査に基づくものである。さらに、21世紀の到来とともに、恐竜の生理機能に関連する二つの重要な特性についての情報が明らかになった。
- 一つは、「恐竜が恒温動物であったか否か」という長年の問題に対する決定的な回答である。
- もう一つは、「恐竜特有の成長速度」についての新たな発見である。
これらの新たなデータは、非常に興味深い展望を示唆している。それに従っていくと、再び恐竜と鳥類の違いという問題に戻ってくる。ここで指摘する鳥類とは、鳥類型恐竜ではなく、現代私たちが認識する全ての特性を備えた「真の」鳥を指す。
なぜ恐竜は存在したのか
なぜ恐竜が存在したのかについて語るため、前章の終わりで触れた三畳紀-ジュラ紀(T-J)境界の大量絶滅から数百万年前まで遡る必要がある。
恐竜が真に地球を支配したのはジュラ紀と白亜紀の期間である。三畳紀においては、彼らはただの普通の小型脊椎動物で、種類も個体数も少なく、低酸素環境で生き残るために努力していた。
しかし、危機の時期は新たな可能性の出現を促す。これは、生命の歴史においてたびたび繰り返される重要なテーマである。多様性は低いままでも、『異質性』[補注]は急速に高まるものである。異質性とは、体制や解剖学的構造の種類の数をいい、恐竜の場合はそれらの差異がとりわけ大きかった。
[補注]「多様性」と「異質性」:「多様性」とは、比較的少数の系統に連なる無数のバリエーションが存在している状態であり、「異質性」とは、根本的に異なる(「異質な」)無数の系統が混在している状態
三畳紀の生物界は低酸素環境のため、喩えて言えば、墜落しつつあるジェット機が死のきりもみ降下のさなかにあるも同然だった。その操縦桿を握っていたのは進化であり、一つの形態を試し、次に別の形態を試し、さらにまた別の形態を試すというような状況であった。
- 恐竜がこの状況から脱出できたのは、前例のないほど『精巧で効率の良い肺』を進化させたからである。
約2億年前、ペルム紀の絶滅からわずか5000万年後、三畳紀は新たな大惨事(ビッグ ファイブの4番目の大量絶滅)で幕を閉じた。
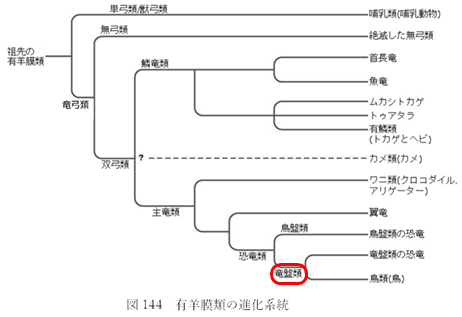
前章で見たように、多くの系統の陸生生物が絶滅の中で、「竜盤類」の恐竜(図144 再掲)だけが無傷で生き残った。
三畳紀の終わりを飾った大量絶滅は陸上だけの現象ではなかった。頭足類のほとんどが絶滅し、生き残ったものがジュラ紀前期に多様化し、「オウムガイ類」、「アンモナイト類」、「鞘形類(現代のイカやタコの祖先)」の三大系統が誕生した。サンゴ礁は再び繁栄し、無数の平たい二枚貝が海底に住むようになった。海生爬虫類の魚竜類と新種の「プレシオサウルス類」が肉食動物の頂点に立った。

陸上では恐竜が繁栄し、哺乳類は小型化し個体数も減少したが、陸上の動物相では目立たない存在であった。しかし、白亜紀の終わり近くには顕著な適応放散が生じ、現代の多くの目が生まれた。鳥類はジュラ紀後半に恐竜から進化した。
これらの事実は既に広く知られているので、これ以上の詳細は省き、代わりにジュラ紀の酸素濃度と、その時代の『ジュラシック・パーク』に存在した恐竜の数と種類との比較に注目したい。
なお、恐竜については、刺激的で興味を引かれるのは、「恐竜はなぜ、どのように絶滅したのか」ではないだろうか。これについては、1980年にカリフォルニア大学バークレー校のアルヴァレスの研究チームが提唱した仮説によれば、6500万年前に小惑星が地球に衝突し、それが環境に影響を及ぼして、かなり急激に白亜紀-第三紀(K-T)境界の大量絶滅が引き起こされた。その際、恐竜が最も目立った被害を受けたために、人々の心に強く焼き付いているのである。
しかし、恐竜に関する疑問を深く掘り下げると、「恐竜はなぜ、どのように絶滅したのか」ではなく、「そもそもなぜ恐竜が誕生したのか」という問いに行き着く。
恐竜が三畳紀中期(約2億3500万年前)に出現したことは確かである。また、これら最初期の恐竜がどのような形状だったかも明確にわかっている。その大部分は、後期の象徴的な恐竜である「ティラノサウルス」や「アロサウルス」を小型にしたような形だった。やはり二足歩行をし、成長が速い。その一方で、一般にはあまり知られておらず、知っていても深く考えたことがない事実もある。それは、2億3000万年前は、「酸素濃度がカンブリア紀以来の最低値に近づいていた時期」だったということである(<物語り その10>図114 再掲)。なぜそこに恐竜が存在したのか、これは我々が解き明かすべき課題である。
現時点でこの謎を解き明かすことが可能である。
動物の歴史を振り返ると、「大気中の酸素濃度と動物の多様性や体の大きさとの間に相関関係が存在する」という事実が繰り返し示されてきた。具体的に言えば、
- 酸素濃度の低い時期は、高い時期より一般的に多様性が低く、体の大きさも小さい。この関係は恐竜についても適用できるようである。
ピーター・ウォードの2006年の著書『恐竜はなぜ鳥に進化したのか - 絶滅も進化も酸素濃度が決めた』(垂水雄二訳、文春文庫)は、恐竜の体制とその後の巨大化が酸素濃度に関連していることを初めて明示した文献である。
恐竜の多様性が大気中の酸素濃度によって決定されると仮定すると、三畳紀に恐竜が出現してから多様性が低い状態が長く続いたことも理解しやすくなる。これは三畳紀後期の酸素濃度が非常に低かったからである。
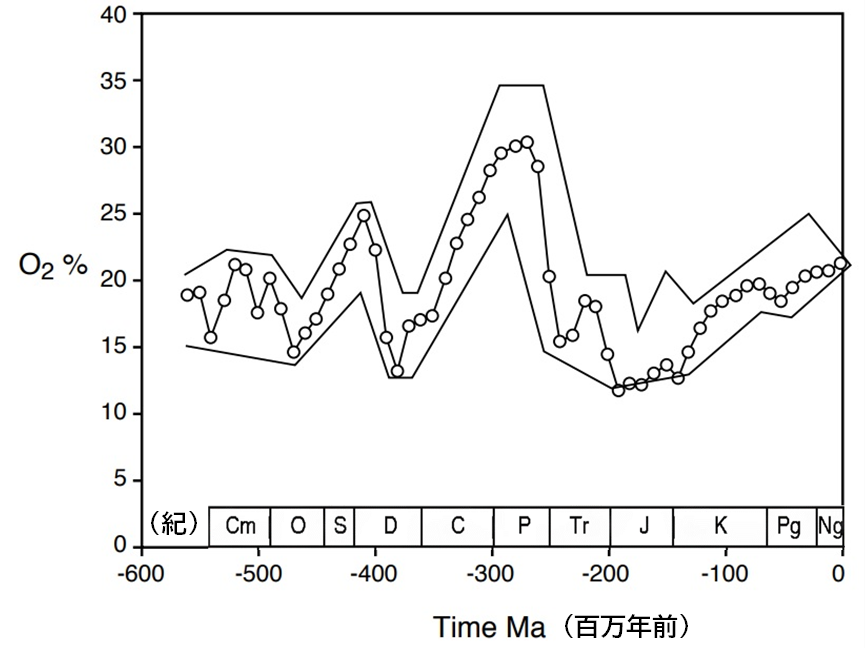
低酸素状況は多くの種を絶滅に追い込みつつ、同時に困難な時代に対応するための新たな生体構造の模索を促進する実験の場となった。これを確認するためには、三畳紀から白亜紀にかけての大気中の酸素濃度に関する最新の推定値と、同じ期間における恐竜の多様性の推移をまとめた資料を比較すればよい。
そのデータは、古生物学者であり堆積学者でもあるデイヴィッド・ファストフスキー率いる研究チームが2004年に発表したものである(注156)。
(注156)David E. Fastovsky, Yifan Huang, Jason Hsu, Jamie Martin-McNaughton, Peter M. Sheehan and David B. Weishampel, Geology 32 (2004) 877–880.
彼らの研究によると、恐竜が初めて登場した三畳紀後期からジュラ紀前期にかけて、恐竜の属の数には大きな変化が見られなかった。しかし、ジュラ紀後期になると属の数が顕著に増加し始め、その傾向は白亜紀末まで続いた。白亜紀後期に入った初期に一度増加が止まったが、白亜紀の終わり(8400万年前~7200万年前のカンパニア期)には、三畳紀からジュラ紀後期までの期間に比べて6~7倍もの恐竜が存在していた。

では、恐竜の属の数がこれほど増加した理由は何か。
酸素濃度と恐竜の属の数の推移を比較すると、酸素濃度が恐竜の多様性を決定づける一因となった可能性があると思われる。
三畳紀後期からジュラ紀前期まで、恐竜の属の数は一貫して少なく、大気中の酸素濃度も現代より低い値が続いた。ジュラ紀に入ると酸素濃度が徐々に高まり、後半には15~20パーセントに達した。そして、属の数が本格的に増え始めるのはそれからであった。酸素濃度は白亜紀を通して着実に上昇し、それに伴って属の数も増加した。白亜紀後期にはその数が大幅に増加し、真の恐竜全盛期を迎えた。酸素濃度が劇的に高まったジュラ紀末には、恐竜の体の大きさも大きくなった。最終的には、既知の恐竜の中で最大級のものがジュラ紀後期から白亜紀にかけて出現した。
もちろん、恐竜が白亜紀に隆盛を極めた理由は他にも存在した。例えば、白亜紀中期に『被子植物』(注157)が出現し、植物界に革命をもたらし、白亜紀の終わり頃には、ジュラ紀に優勢だった針葉樹の大部分が顕花植物に取って代わられていた。被子植物の台頭により、植物の種類が増え、それが昆虫の多様化を促進した。
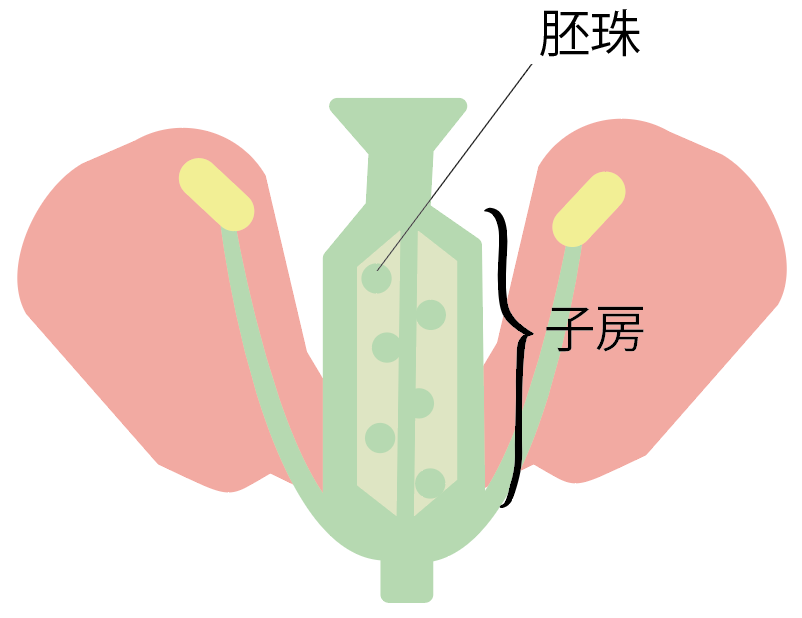
(注157)『種子植物(顕花植物)』(花をつけ種子を作る植物)のうち、一般に花と呼ばれる生殖器官の特殊化が進んで、胚珠が心皮にくるまれて子房の中に収まったものをいう。心皮が発育して果実となる。もう一つの分類群は『裸子植物』と言われ、これは胚珠が子房にくるまれておらずむき出しになっており、果実も作らない。
利用できる資源が増えたことも多様化の引き金となったかもしれない。だが、酸素と多様性の関係、そして酸素と体の大きさの関係は、様々な種類の動物を通して繰り返し確認されている。昆虫や魚類のほか、爬虫類や哺乳類でも同様の関係が見られる。恐竜がその例外とは考えにくい。
恐竜が出現したのは、三畳紀後半の低酸素期(酸素濃度は10~12パーセントで、現在の標高4500メートル地点に相当)の最中、あるいはその直前だった。その時点での酸素濃度は過去5億年間で最も低かった。前述したように、他の動物種は酸素濃度の極端な変動に応じて体制を変化させることが多かった。恐竜も同じであった。恐竜の体制は初期の爬虫類とは根本的に異なっていた。それが現れたときには酸素が最低値を記録し、しかも地球規模での酷暑のさなかだった。これが偶然だとは断言できない。
しかし、「恐竜的特徴」を多数生み出す要素が、低酸素環境への適応から導かれると考えると、単なる偶然であったとは考え難い。初期恐竜(スタウリコサウルスやその後に出現したヘレラサウルス等)の体制を観察すると、その時代の低酸素環境に応じて生まれた特徴が見受けられる。この観察から、


初期の恐竜の『二足歩行体制』は、三畳紀中期の低酸素環境への反応として進化したと結論づけることができる。
- 最初期の恐竜は二足歩行の姿勢をとることで、『キャリアの制約』による呼吸の制約を克服したのである。
- つまり、三畳紀の低酸素環境が触発要因となり、この新体制が形成され、結果として恐竜が誕生したのである。
三畳紀の終わりの2000万年間、海抜ゼロメートルでも酸素濃度が10パーセントしかなかったと推定されている。そのような環境での生存については、まだ十分に理解されていない。現代において、同等の酸素濃度の場所は、ワシントン州のレーニア山山頂(標高4392メートル)である。これはまた、ハワイ最高峰の火山の頂上と同じ酸素量であり、そこでは大型のケック天文台が宇宙を観測している。ここで観測を行う天文学者が即座に感じることは、酸素濃度が非常に低いためエネルギーが消耗し、思考が遅くなることである。
だが、ここでは「斉一説の原理」(条件に変化がなければ自然現象は同じように繰り返されると仮定すること)は役に立たない。それは、標高の高い場所を参照にして低酸素環境を理解しようとすると、多くの問題が生じるからである。標高が上がると酸素だけでなくすべての気体の濃度が薄くなるためであり、その一つである水蒸気は高地において鳥の卵に有意な影響を与えている。
それでも、当時の陸上生物が進化する上で最大の制約は「低酸素」であったという事実は変わらず、それに対応するために大規模な適応構造を発展させていたはずである。 そして実際にその通りだった。私たちはそうした動物の一つを『恐竜』と名づけた。
- 初期の恐竜は全て二足で歩き、新種の肺を使用して新たな方法で呼吸した。
- 結果として恐竜は、低酸素環境において史上最も効率的に活動できる陸上生物に進化した。
- 鳥類はその進化の生き残りであり、同じ利点を保持し続けている。

化石記録によると、「本物の恐竜」と呼べる最初期の動物は二足歩行であり、その化石の年代より少し早い三畳紀のどこかの時点で『主竜形類』(224頁「三畳紀における生物の復活」節を参照のこと)から進化した。主竜形類は恐竜より原始的な二足動物であり、後にワニ類の祖先となる系統で、恒温性だったか、あるいは恒温性へ進化する途中であった可能性がある。二足歩行は、このグループで反復して現われる形質であり、初期の段階では二足歩行のワニ類まで存在した。
では、なぜ二足歩行が低酸素への適応となったのか、その理由を考えよう。
現代でもトカゲは走りながら呼吸することができない。これは、腹這いでのたくるように進むためである。現代の哺乳類は、独特のリズムで四肢の動きと呼吸を同時に行っている。たとえば、ウマやジャックウサギ、チーターなどは、一歩進むごとに一呼吸する。いずれも四肢は胴体から真下に伸びて、それを可能にするために、腹這いの爬虫類よりも背骨が格段に頑丈である。哺乳類が走るときには、背骨はわずかに下向きに湾曲したり、完全に直線に伸びたりする。このわずかな上下の動きが、息を吸い込む動作や吐く動作と連動している。しかし、この呼吸法は、三畳紀に真の哺乳類が登場するまで存在しなかった。三畳紀で最も進化していたキノドン類でも、まだ体全体が地面から完全には離れていなかったため、走行中の呼吸は苦労したはずである。しかし、
- 四本ではなく二本の脚で走れば、肺や胸郭への影響は少ない。呼吸と運動が分離できるため、獲物を高速で追い詰める際でも、必要な回数だけ呼吸が可能である。
酸素濃度は低かったが、捕食が活発だった時代には、獲物を狩るためや捕食者から逃れるために、少しでも有利な特性があれば、生存の可能性は確実に高まる。その有利な特性が、食物を探すための時間や方法を改善するだけでも良い結果をもたらす。ペルム紀後期に活躍した腹這いの捕食者(例えば、獰猛なゴルゴノプス類など)は、待ち伏せ型の狩猟を行っていた。その時代もそれ以前も大半の捕食者はそうだったし、今日のトカゲ類もすべて待ち伏せ型である。能動的に獲物を追い詰めるタイプの捕食者にとっては、素早さと持久力が必要であった。
ワニの系統と恐竜の系統は三畳紀に起源を持ち、双方は共通の四肢動物を祖先としているとされる。この祖先は南アフリカ由来の爬虫類、「ユーパルケリア」であると推定されている。このグループは『鳥頸類』(注158)という専門用語で表され、その最初の種から『二足歩行』へ向けた進化が開始された。
(注158)「鳥頸類」は、『主竜類』を形成する主要な系統群であり、鳥類に繋がるグループである。このグループは恐竜とそれに近縁な分類群から構成され、さらに『恐竜様類』と『翼竜類』に大別される。長い足と独特な足関節、そして細長い頸部を特徴とする。
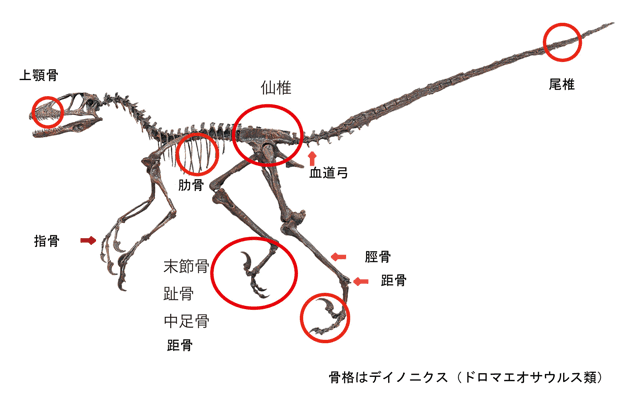
進化の証明となるのは「距骨」(くるぶしにある骨)である。四肢動物の場合、その他の同じ位置に存在する骨とは異なり、距骨は簡略化され、単純な蝶番関節になっている。この特徴に加えて、前肢より後肢が長く、首が伸びて緩いS字を形成することも二足歩行の証拠とされている。こうした初期の「鳥頸類」が二つの別々の系統に分岐し、一方は空を飛ぶ能力を獲得した。この空飛ぶ系統が『翼竜』である。三畳紀後期の鳥頸類であるスクレロモクルスが、翼竜の最初の種だった可能性が示唆されている。この種はまだ陸上動物の形態を保持しており、走行速度が速いように観察される。おそらくは腕についた皮膚のたるみを利用して、大股の歩行の間に滑空するように進化したと考えられている。

間違いなく空を飛んだ最初の翼竜は、やはり三畳紀後期の『エウディモルフォドン』である。この翼竜系統の鳥頸類が空を飛ぶ進化を遂げた一方で、陸生の姉妹グループは初期の恐竜の形態へと進化していった。三畳紀の『ラゴスクス』は過渡的な形態を持ち、二足歩行動物と四肢動物の中間的な存在である。遅い速度で移動するときには四肢をすべて使用するが、勢いよく走るときには後肢で立ち上がると推定される。ラゴスクスは捕食者であり、獲物を追い詰めるために高速で走行する必要があった。しかし、その前肢と手の形態が恐竜型と呼べる段階に達していなかったため、ラゴスクスは恐竜とは認定されていない。
その後継種である三畳紀の『ヘレラサウルス』(図178参照)は、全ての条件を満たしているため「恐竜」に分類されている。
- これが恐竜第一号である。
- ただし、ヘレラサウルスには一つ欠けていた特性が存在し、その後の直接の子孫になってからその点が改良されたと見られている。それは、大気中酸素濃度の低下に対応するための『新しい種類の呼吸システム』である。
この最初の恐竜は完全に二足歩行で、両手で物を掴む能力を持ち、私たちと同じような親指を有していた。手には五本の指があり、三本指の足とは全く異なる形状をしていた。実際には足も五本指だったが、二本は退化しており、歩行や走行時には地面に触れるのは三本だけであった。既に四足歩行ではないため、地面に到達するような手を維持する必要性は進化の観点から見て存在しない。では、移動に不要な余剰の肢はどのように使用されたのだろうか。後世に出現する著名な『ティラノサウルス・レックス』は、前肢が非常に小さいため、その肢が機能していなかった考えられる一方で、恐竜第一号の「ヘレラサウルス」は異なる。姿勢を見ると、後年の肉食恐竜と同じだが、手は明らかに使用されていた。おそらく、走行しながら手で獲物を捕まえる、もしくは抱えるなどしていたと推測される。
これらの特性は、その後の全ての恐竜の起源となる体制を形成していた。その特性は次の通りである。
- 二足歩行
- 長く伸びた首
- 物を掴むことのできる手
- 実際に機能する親指
- さらには骨盤も大きくて特徴的だった。これは、骨盤の表面積を広くして大量の筋肉を付着させ、その筋肉を使って歩いたり走ったりしていたからである。
この初期の二足歩行恐竜は比較的小型で、三畳紀が終わる前に二つのグループに分岐した(図185参照)。それが恐竜を区別する最も根本的な分類として残った。
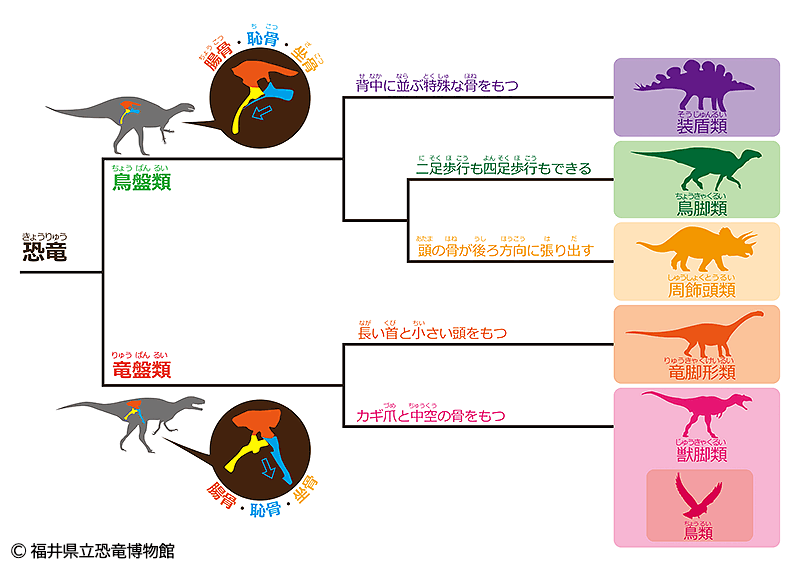
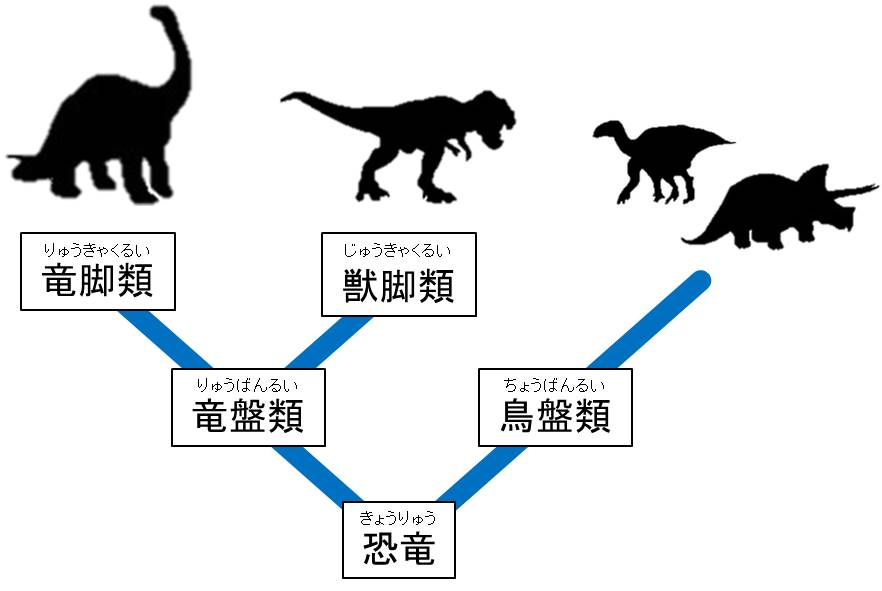
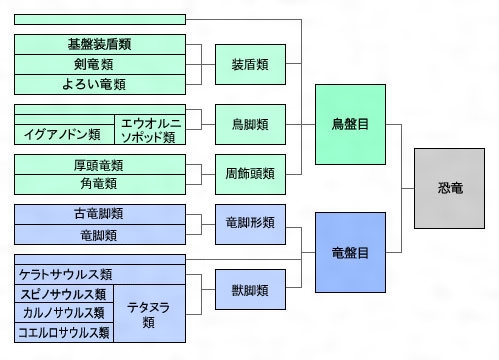
具体的には、三畳紀の二足歩行恐竜のうち一種が『寛骨』(注159)の形を変え、初期の恐竜では前向きだった恥骨を後ろ向きに変化させたのである。そして、この変化を境にして恐竜は大きく二つに分かれた。一方は祖先の構造を継承した『竜盤類』(注160)、もう一方はその子孫である『鳥盤類』(注160)となり、その後約1億7000万年間にわたって両者が世界を支配することになる。
(注159)骨盤部分を構成する左右一対の不規則形の骨で、「腸骨」、「坐骨」および「恥骨」が癒合してできた骨
(注160)恐竜は、「骨盤」の特徴から大きく『鳥盤類』と『竜盤類』の2つのグループに分けられている。それぞれ、現在の鳥の骨盤、トカゲの骨盤に形が似ているということで名付けられた。「鳥盤類」はさらに、ステゴサウルスなどを含む『装盾類(そうじゅんるい)』、イグアノドンなどを含む『鳥脚類(ちょうきゃくるい)』、トリケラトプスなどを含む『周飾頭類(しゅうしょくとうるい)』に分けられる。また「竜盤類」は、ブラキオサウルスなどを含む『竜脚形類(りゅうきゃくけいるい)』とティラノサウルスなどを含む『獣脚類(じゅうきゃくるい)』に分けられる。これらのグループもそれぞれがさらに細かく分類されている。
恐竜の呼吸器系は、現代に存在する変温動物であるトカゲのものとは全く異なり、恒温動物である鳥類のものに似ていたことが明らかになっている。現代の『羊膜類(爬虫類、鳥類、哺乳類)』の肺には二つの基本形が存在する。羊膜類の祖先である石炭紀の爬虫類は、肺が単純な袋状であったため、そこから二種類どちらのタイプの肺が発生しても不思議ではないであった。
一つは『肺胞式』の肺で、現生哺乳類のすべてがこれを有している。もう一つは『隔壁式』の肺で、現存する爬虫類と鳥類がこのタイプである(<物語り その13>図172参照)。
- 肺胞式の肺は、肺胞と呼ばれる球状の袋が多数集まって形成され、肺胞には血管が張り巡らされている。空気は肺胞を出入りするので、双方向のシステムといえる。哺乳類はこの『肺胞式』である。
私たちが行っている「吸って、吐いて、吸って、吐いて」という呼吸法は、肺胞式に典型的なものである。吸い込んだ空気は肺の中の袋に引き込まれ、その後再び吐き出され、その過程で酸素が二酸化炭素に交換される。これを行なうためには、筋肉の力で胸郭を拡張する動作と、横隔膜と呼ばれる大きな膜状の筋肉を収縮する動作を組み合わせる。直感的には不思議な気がするが、横隔膜が収縮すると肺の容積は増大する。この二つの動作によって肺内の気圧が低下し、空気が流れ込む仕組みとなる。息を吐くことは、ある程度は各肺胞の弾性反発によって行われる。肺胞は膨らんで拡大し、その弾性特性によって自然に収縮する。この形式の肺には多くの肺胞が使用され、そのおかげで効率よく酸素を獲得することが可能である。私たちは、動きに満ちた活発な生活様式を維持するために、この仕組みが必要である。しかし、空気の出入りに同じ管を使用することは無駄を生じやすく、しかも酸素を取り込むのに費やすエネルギー量の割には酸素摂取量が少ないという欠点がある。
- これに対して、爬虫類や鳥類の肺は『隔壁式』で、いわば一つの巨大な肺胞のようなものと考えればいい。それをより小さな空間に仕切って気体交換する表面積を増やすために、葉のような細胞組織のシートが多数、肺の中に広がっている。このシートのことを『隔壁』というので、このタイプの肺は「隔壁式」と呼ばれる。
これが基本デザインであるが、動物の種類によって様々な変型が見られる。小さな部屋に区切られているものもあれば、第二の袋が肺の外側にあって、管でつながっているものもある。肺胞式の肺と同様に空気の流れは通常双方向であるが、すべてがそうではない。近年その例外が見つかったことで、初期爬虫類の生物学的特性はもちろん、ペルム紀大量絶滅における爬虫類の運命についても私たちの理解は根本的に変わった。
隔壁式の肺には弾性がないので、息を吸い込んだ後で自然に収縮することはない。肺の換気方法は動物のグループによって異なる。トカゲ類とヘビ類は肋骨の動きを利用して空気を吸い込むが、すでに見たようにトカゲ特有の歩き方のため、肺の空洞を完全に広げることができない。そのため、トカゲは移動中に呼吸ができない。
最も非効率なのは哺乳類の呼吸法、つまり、吸気と呼気がどちらも同じ管を通るシステムである。非効率な理由は、息を吐き出して次に吸うときに気体の分子が入り乱れるからである。呼吸が速い場合、空気の吸入が始まる前に呼気が外に出ようとして、無秩序な衝突が生じてしまう。その結果、吐き出すべき気体、つまり二酸化炭素が多くて酸素が少ない空気が、再び吸い込まれることが頻繁に起こる。
長らくワニ類も同じ問題を持っていると考えられてきた。しかし、2010年の研究で、実はワニ類が呼気と吸気それぞれの一方通行経路を使っていることが明らかになった(<物語り その13>注154参照)。これは鳥類や恐竜と同様である。また、ペルム紀から三畳紀にかけて生息した古代の爬虫類の祖先も、同時代の獣弓類(哺乳類の祖先)より呼吸の効率が高かったことが示唆されている。
爬虫類の祖先からは、最終的に現代のワニ類や鳥類、そして絶滅した恐竜が誕生している。これらの動物がペルム紀絶滅というフィルターを突破できたのは、生き残るための二つの大きな利点を備えていたからである。
- 一つは「変温動物」だったこと、
- もう一つは、「哺乳類や哺乳類型爬虫類よりも空気中から多量の酸素を取り込む能力を持っていた」こと
であった。
このような状況は私たち哺乳類にとって不利であった。大量絶滅の危機と混乱の中、競争が激化し、私たちは勝算を見つけられなかったのである。これは単に生き延びるためだけでなく、最終的に優位にたつための競争でもあった。結果として、哺乳類は中生代を通じておおむねネズミ程度の大きさのままで過ごすことになった。おそらくは恐竜に囲まれ、ネズミのように恐怖に陥った状態だったであろう。
鳥類の気嚢システム
陸生脊椎動物の肺には、隔壁式の肺の一変種が存在している。その最たる例が鳥類の肺と呼吸器系である。
鳥類の肺自体は小さく、柔軟性に欠けるため、私たちの肺のように一回の呼吸で大きく膨らんだり縮んだりはしない。その代わり、胸郭が呼吸に深く関与している。特に骨盤領域に最も近い肋骨は、胸骨の底部に接続する部分で可動性が非常に大きく、この可動性が非常に重要な役割を果たしている。しかし、
最大の違いは、現存する爬虫類や哺乳類とは異なり、鳥類の肺には『気嚢』と呼ばれる付属器官が存在し、結果的に非常に効率的な呼吸が可能になっていることである。すなわち、私たち哺乳類(および鳥類以外のすべての生物)の肺は袋状であり、空気を吸い込んでから吐きだす仕組みとなっているが、鳥類はまったく別のシステムを持っているのである。
図172(<物語り その13>)に示すように、鳥が空気を吸い込むと、空気はまず気嚢群に入り、その後肺本体の組織に流れ込む。この際、空気は気管からではなく接続している気嚢から流れてくるため、肺の中では空気が一方向に流れる。そして最後に肺から呼気が出てくる。
このように肺細胞膜を通過する空気の流れが一方通行であるため、『対向流系』を形成できるのである。
- これは、「空気が一方向に流れ、血液が肺内部の血管を空気と反対方向に流れる構造」を指す。
- 対向流を利用したガス交換は、袋状の肺よりも酸素をより効率的に取り込み、二酸化炭素を放出できるのである。
『気嚢式』が哺乳類のシステムより優れていることは明らかである。海抜ゼロメートルでの空気中からの酸素摂取効率は、鳥類が哺乳類より33パーセント高いと推定されている。しかし、高地ではこの差がさらに大きくなる。地上から約1500メートルの高度での酸素摂取効率は、鳥類の方が200パーセント高いのである。このため、高地では哺乳類や爬虫類と比べて鳥類が圧倒的に優位に立つことが可能である。遠い過去には、海抜ゼロメートルでの酸素濃度が、現在の標高1500メートル地点よりも低い時期が存在した。そのような時代に鳥類型の呼吸システムがあれば、非常に大きな利点となっただろう。なぜなら、競争相手も捕食相手もそのようなシステムを持っていなかったからである。
現在では、鳥類が小型の二足歩行恐竜から進化したことが明らかになっている。この恐竜は、最初期の恐竜である『竜盤類』の系統に属していた。最初の鳥類の骨格はジュラ紀の地層から発見されたが、鳥類の肺に付属している気嚢は軟組織であるため、特殊な環境下でしか化石化しない。このため、気嚢式の肺がいつ誕生したかを直接的に知る手段は存在しない。
しかし、間接的な証拠なら存在し、その証拠には十分な説得力がある。そのため、
- 恐竜が気嚢を持っていたと考える研究者たちは、「すべての竜盤類に現代の鳥類と同じ気嚢システムが存在していた」と結論づけているのである。
- さらに、「恐竜も鳥類と同様に恒温性であった」という新たな説が提唱された。
- この説の根拠は、恐竜の骨に見られる空洞であり、これが気嚢の存在を示していると考えられている(注161)。
2002年に恐竜学者グレッグ・ポールが提唱した説によると、『主竜類』と呼ばれる最初の生物群が既に気嚢を持っていたとされる。これらの『主竜類』は、ペルム紀後半から三畳紀前半にかけて生息した原始的な爬虫類であり、彼らからは最終的にワニ類や恐竜類、鳥類が進化した。
(注161)哺乳類が肺の拡張や横隔膜収縮、すなわち胸郭周囲の筋肉による呼吸を発達させたのに対して、鳥類は気道に『気嚢』というポンプを設置し、空気の吸い込みと押し出しを複数のポンプで効率化したことが知られている。鳥類の祖先である「竜盤類」も「気嚢システム」を持っていたと推測されており、それを裏付ける証拠も存在する。
鳥類の直接の祖先である獣脚類はもちろん、巨大な種が多かった竜脚類も体温の過剰上昇を防ぐラジエーターとして、あるいは重い骨を軽減する空洞として、「気嚢システム」が必要だったと考えられている。飛行する鳥類は体重を軽減するために骨を空洞化し、気嚢システムの一部として活用している。鳥類の呼吸システムは哺乳類よりも優れており、標高1500メートル(ほぼジュラ紀の酸素濃度)でも酸素吸収効率はほぼ2倍である。
21世紀の初めにはまだ明らかになっていなかったが、前述の通り(<物語り その13>図172参照)、ワニ類とその近縁種の呼吸器系は、当時の定説よりも優れていた。その理由は、「呼吸器系の中で空気の流れを一方通行にする」という革新的な仕組みにあった。この事実は2010年になってようやく明らかになった。
この発見により、ワニ類、恐竜類、哺乳類の中で生存能力が最も高かったのはどれか、という視点が大きく変化した。特に、三畳紀の爬虫類はすべて、私たち哺乳類よりも優れた呼吸能力を持っていたと考えられている。しかし、その後、気嚢システムが急速に進化したのは、少なくとも恐竜につながる系統であったと見られている。一方、ワニ類は、新たな呼吸器系を獲得したにも関わらず、大きな改変を加えることを止めてしまった。すなわち、含気骨や気嚢の導入を試みることはなかった。
最初の真の恐竜が三畳紀中期に出現した時点で、気嚢システムは既に部分的に発達していたと推察される。最初の恐竜である最も原始的な『獣脚類』には含気骨が見られないが、肺本体は比較的小さく、柔軟性に欠けていたと推察される。これらの特徴は現生の鳥類の肺と一致する。
ジュラ紀のアロサウルスなどの恐竜では、気嚢システムがほぼ完成していた可能性がある。しかし、その形状は飛行のために改良を重ね、胸部と腹部に複数の気嚢を有する鳥類のものとは大きく異なっていた。
ジュラ紀中期に『始祖鳥』が出現した頃には、恐竜の呼吸器系には様々なタイプが存在したと推測される。「含気骨」を持つ種と持たない種が存在したであろうし、「収斂進化」(注162)も進んでいたと考えられる。例えば、ウェーデルが調査した結果(注163)によれば、大型獣脚類では骨の含気化が広範囲に及んでおり、これは二足歩行の竜盤類の気嚢システムとは無関係に独立して生じた可能性がある。
(注162)異なる系統の生物が、環境要因などで同様の選択圧に曝されることにより、似通った形態へとそれぞれ進化を遂げるような現象。
(注163)大きな竜脚類の椎骨は、『含気骨』という、空気で満たされた穴だらけの骨だった。カリフォルニア州ウエスタン健康科学大学の古生物学者マシュー・ウェーデルは、「竜脚類の頸椎の質量は、通常の骨の充填物質の35%ほどしかなく、そのおかげで首を最大15mまで伸ばすことが可能であった」と述べている。含気骨内の空洞部分は体腔の気嚢に接続されていたと推測される。気嚢は、現生鳥類の特徴であり、肺に空気を効率よく送り込む能力を持つ。こうしたメカニズムにより、巨大な恐竜の呼吸効率が向上した。気嚢による換気がなければ、竜脚類は1回の呼吸で首を満たす排気を排出することができなかっただろう。肺だけでは体積が不足していたのである。
この事実は、含気椎骨が巨大な体と関係した適応だと解釈するかもしれないが、ウェーデルは初期の小型の竜脚形類である『パンティドラコ』に、ある機能の先駆けと見える特徴を発見した。その頸椎には凹みが存在し、それは竜脚類の頸椎の穴の位置と一致していた。したがって、小型の恐竜において、原始的な気嚢と含気骨はどのように役立ったのだろうか。研究者らは、これらが酸素交換の効率を向上させたという仮説を立てている。この仮説が正しければ、恐竜の祖先は、大気中の酸素濃度が現代よりもはるかに低かったペルム紀後期から三畳紀前期(約2億6000万~2億4000万年前)において、他の生物種との生存競争に勝利できた可能性がある。
なお、気嚢は「竜盤類」恐竜全般に見られるが、他方、「鳥盤類」の大型恐竜(『ハドロサウルス科』の恐竜や『イグアノドン類』、角のある『角竜類』恐竜)はジュラ紀ではなく白亜紀の恐竜なのは、偶然ではない。「鳥盤類」に気嚢システムがない理由は、これらが生息した時代を考えればよくわかる。酸素濃度が非常に低かったジュラ紀のあいだ、鳥盤類はごく小さな勢力にすぎなかった。ジュラ紀後期から白亜紀にかけて酸素濃度が大きく上昇してようやく、この第二の恐竜グループが広く分布するようになったのである。


ジュラ紀の時代には恐竜だけでなく、私たちの直接の祖先も小型ながら存在し、陸上・海洋生物の中にはリクガメやウミガメ、首の長いプレシオサウルスやワニなども含まれていた。それでも、陸上を支配していたのは間違いなく恐竜であった。
一見すると恐竜の体型は多種多様であるように見えるが、実際には基本的な型は三つしか存在しない。
- 三つとも鳥類や哺乳類と共通する特徴をもっていた。つまり体が完全に起きていることである。
- 具体的には、図191に示すように、『二足歩行型』と、『首の短い四足歩行型』、そして『首の長い四足歩行型』である。
これらの型が誕生した時期や、最も個体数が多かった時期はそれぞれ異なる。恐竜の『モルフォタイプ(体制)』の変遷は、それぞれの生息した時代に応じて以下の五段階に分類することができる。


- 三畳紀後期 最初の恐竜は三畳紀後期に出現したが、その後1500万年間、多様性は低いままであった。大部分を占めたのは二足歩行で肉食性の『竜盤類』であった。三畳紀の終わりにかけては、四足歩行の竜盤類が現れた。これらを『竜脚類』と呼ぶ。『鳥盤類』は三畳紀が終わる前に竜盤類から分岐したが、恐竜全体の種数や個体数に占める割合はごく僅かであった。三畳紀を通じて、恐竜の体長は1~3メートル程度と小さかった。
- ジュラ紀初期~中期 この期間、「竜盤類」の二足歩行恐竜と首の長い四足歩行恐竜が動物界を支配していた。一方で、「鳥盤類」は小型で少数のままながらもいくつかの主要な祖先系統へと分岐した。最終的にこれらの系統は白亜紀に入り、恐竜の中で最も多様化した。しかし、最も顕著に多様化したのは、竜盤類の一つである「竜脚類」であった。竜脚類は三畳紀後期に、「古竜脚類」と「真の竜脚類」の二系統に分岐した。なお、『古竜脚類』は、アンキサウルス、プラテオサウルスなどに代表される草食恐竜の分類群である。『真の竜脚類』は、サルタサウルスなどヴルカノドンのような原始的な竜脚類に比べてよりティタノサウルスに近縁である全ての竜脚類で構成される竜脚類恐竜の分岐群である。
二足歩行の竜盤類も多様化し、ジュラ紀の初期と中期に繁栄した。これらは三畳紀末期に二つのグループ(ケラトサウルス類とテタヌラ類)に分岐していたが、テタヌラ類はさらに三つに分岐し、スピノサウルス類、カルノサウルス類、そしてコエルロサウルス類となった。
コエルロサウルス類からは最終的に、史上最も有名な恐竜である白亜紀の『ティラノサウルス・レックス』が進化した。ただし、ジュラ紀中期の時点ではまだ小型であった。ジュラ紀で最も重要な進展は、『鳥類へとつながる祖先系統』が誕生したことであった。 - ジュラ紀後期 『巨大恐竜』の時代である。史上最大の竜脚類の化石はこの時期の岩石中から見つかっており、これらの生物による支配は白亜紀の初期まで持続した。竜脚類と同じような大きさを誇るのが竜盤類の巨大肉食恐竜で、その代表例として獣脚類の『アロサウルス』(図186参照)が存在した。したがって、この時代の最も注目すべき特徴は、ジュラ紀初期や中期よりも大幅に大型化した恐竜の出現である。これは竜盤類に限らず、装甲を有する鳥盤類もこの時期に大型化した。とりわけ大型化が顕著だったのは、頑丈な装甲をもつ『ステゴサウルス』であった。「ステゴサウルス類」、「アンキロサウルス類」、「ノドサウルス類」、「カンプトサウルス類」、「ヒプシロフォドン類」の出現により、鳥盤類は多様化し、恐竜類の外観は大きく変化した。
- 白亜紀初期~中期 当初は大型竜脚類による支配が続いたが、白亜紀が進むにつれて大きな変化が発生した。鳥盤類は更に多様化し、その個体数も増加した結果、竜盤類を上回った。ジュラ紀末に竜脚類の属が数多く絶滅したことにより、竜脚類の存在数はますます減少した。
- 白亜紀後期 恐竜が急速に多様化した。この多様化の中心となったのは、数多くの新種鳥盤類で、特に「ケラトプス類」、「ハドロサウルス類」、「アンキロサウルス類」が顕著であった。竜脚類はごく一部の個体しか生息していなかった。


進化の歴史が一つの要因だけで決定されることはない。恐竜の形態変化も同様である。捕食者と被食者の相互作用、恐竜同士や同時代の他の動物との競争が影響を及ぼしたこともあれば、気候変動が一部の原因となった可能性も存在する。この気候変動を引き起こしたのは、ジュラ紀から白亜紀にかけての海面の大きな上下動であった。さまざまな要因が考えられる中で、『酸素濃度』も一役買っていたと推測される。
我々の分類では、三畳紀後期は第一段階に位置し、これは低酸素の時代であった(<物語り その10>図114参照)。加えて、二酸化炭素濃度が極めて高かった(<物語り その13>図170参照)ことが、小惑星の衝突ではなく、三畳紀-ジュラ紀(T-J)境界大量絶滅の主な原因であった。低酸素と全球的な高温の組み合わせは、生物の死を引き起こすメカニズムとなった。
しかしながら、この大量絶滅の前後に生息していた陸生脊椎動物の分類群の数を調査した結果、竜盤類恐竜が最もうまく大量絶滅を生き延びたことが明らかとなった。
- その重要な理由の一つは、『優れた呼吸器系』ではなかったかと考えられる。すなわち、『気嚢式の肺』を持っていたことが、他の肺を持つ陸上動物との生存競争において優位に立つ要因となったのである(<物語り その13>注154参照)。
一方の鳥盤類恐竜には、竜盤類のような効率的な呼吸器系は持っていなかった。しかし、竜盤類より頭部が大きく、顎が強く、歯も優れていたため、食物の獲得においては有利であった。白亜紀に入って酸素濃度が現代レベル近くまで上昇すると、その利点が幸いして鳥盤類が主要な植物食恐竜となり、『競争的排除』(食物・生息場所などの生活要求が類似した種は、競争の結果として同じ場所に共存できないこと)によって多くの竜盤類の植物食恐竜が絶滅に追い込まれた。
ジュラ紀から白亜紀にかけて、大気中の酸素濃度は比較的急激で大幅に上昇していたが、同時に他のいくつかの重要なイベントも発生していた。
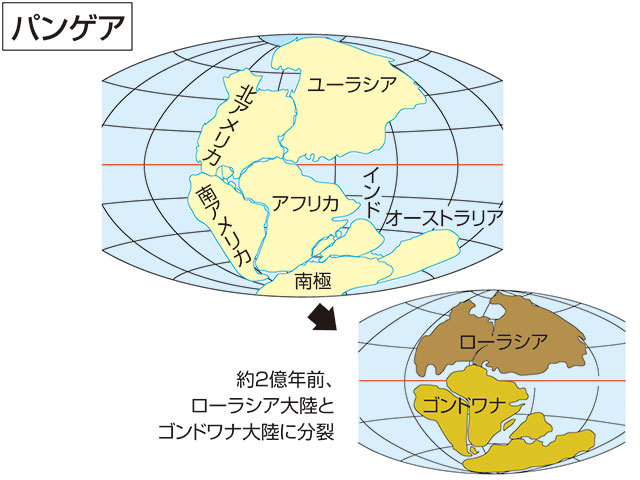
- その一つは、それまで地球上唯一の超大陸であった『パンゲア』が、より小さな大陸群に分裂したことである。
- また、中生代後期の恐竜類の分布や分類学上の構成に大きな影響を与えた出来事もあった。それは植物相が根本的に変化したことである。
恐竜は裸子植物(針葉樹類、シダ種子類、ソテツ類、イチョウ類など)が支配する世界で進化した。しかし、白亜紀の初期には新たな種類の植物が出現した。「花を咲かせる植物」である。これらは『被子植物』(注164)と呼ばれ、新しい生殖方法と様々な適応構造を備えていたため、急速な適応放散を遂げた。最終的には、地球のほぼ全域で古い植物相との競争に勝利し、約6500万年前の白亜紀末には植物全体の九割を占めるまでに至った。
(注164)『裸子植物』は、将来種子になる「胚珠」が裸で露出している植物のことを指し、『被子植物』は、将来種子になる「胚珠」が子房によって覆われ隠れている植物のことである。
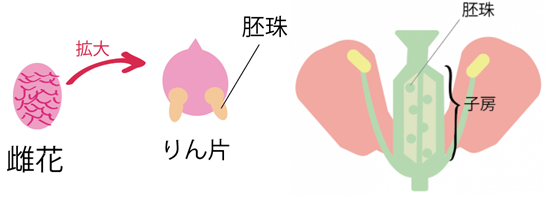
植物食動物にとって食物の種類が変化すると、その影響を避けることは難しいだろう。同様に、肉食動物も、自身が獲物とする植物食動物の種類が変わると、その変化が体制に直接的な影響を与えることになるであろう。
植物食で生き抜くためには、食糧となる植物に適した歯が必要である。竜脚類はマツ類の針状葉を食べて生活し、その巨大な樽状の体を発酵タンクのように利用し、消化が難しい食糧源を処理していたと考えられている。被子植物である広葉樹が出現すると、歯の形状や咀嚼面は、針状葉を木から剥ぎ取るものとは異なるものが必要になったはずである。
したがって、
- ジュラ紀に竜脚類が支配していた動物相が、白亜紀に鳥盤類が支配する動物相へと変化した理由の一部は、植生の変化によるものであったと考えられる。
それにもかかわらず、
- 呼吸にも関連性があった可能性がある。
- そしておそらく、酸素濃度が15パーセントを超えなければ、鳥盤類が覇権を握ることはできなかったであろう。
三畳紀からジュラ紀の恐竜の肺と鳥類の進化
最初の恐竜は、それまで存在していた動物とも、現在生息している動物とも大きく異なっていたとされている。
「直立した姿勢」と、「進化しつつある気嚢システム」を通じて、同時代の他の動物よりも『呼吸効率』(単位時間当たり、もしくは呼吸に消費される単位エネルギー当たりに大気から取り込む酸素の量)が高かったと考えられている。
- しかし、初期の恐竜は『内温性(体温が主に代謝熱で維持されている状態)』を失い、その代わりにより受動的な『恒温性(恒温性とは内温性のうち、自律的に体温を制御している状態)』を獲得した可能性がある。
- その結果、恐竜は安静時に恒温性を活用して酸素の消費を減らし、活動時には高度な肺システムを使用して長時間の運動を無酸素状態に陥ることなく続けられたのである。
- 鳥類は恐竜の一グループであり、ジュラ紀に登場した。そして最終的には、「現存の爬虫類とはまったく異なる肺システム」と、「内温性」([補注] 鳥類は羽毛をもち空を飛ぶ内温性(体温が体内で生ずる代謝熱によって維持される性質。定温動物 以外にも、大型の爬虫類、回遊魚、昆虫の一部は内温性である)の陸上脊椎動物である。)の両方を獲得したことがわかっている。
鳥類型の恐竜と恐竜型の鳥類
特別な関心が集まる「ティラノサウルス類」を除き、近年最も注目されている恐竜のグループはおそらく「基盤的鳥類」である。議論の焦点は主にその外見についてであるが、何より重要なのは飛び始めた時期とその理由である。

最初の鳥類は約1億5000万年前に出現した。最初の鳥類として最も有名なのは依然として『始祖鳥』である。
- この時期はジュラ紀が始まる直前で、すでに5000万年以上にわたって酸素濃度が上昇していた。
- 当時の恐竜は巨大なものが多く、鳥類の直接的な祖先は、地上を素早く走る恐竜で、捕食のために前肢を使用していたと推測されている。カリフォルニア大学バークレー校の古生物学者ケヴィン・パディアンによれば、その動きは飛行動物の羽ばたきの前適応であった。
- 化石記録からは、鳥類の祖先が二足歩行の肉食竜盤類であったことが示唆されている。具体的には、『トロオドン類』か『ドロマエオサウルス類』で、どちらもすでに羽毛を生やしていたと考えられている。


始祖鳥が飛べたのかどうかについては、今日ではほとんどの専門家が肯定的であるものの、本格的な飛行が始まった時期については意見が分かれている。
ジュラ紀後期の空には多種多様な「プテロダクティルス類」(翼竜の一種)が繁栄していたが、その当時の『鳥類』が実際に飛べたのかは疑問である。

化石記録を見ると、白亜紀前期にはすでに鳥類(エオアルラヴィス)が存在し、「親指からの翼」が発展していた。これは機動性を高め、より低速の飛行を可能にする適応である。したがって、
始祖鳥の登場から数百万年のうちに、飛行はかなりの進歩を遂げていた。中国での新たな発見により、白亜紀の初期にはすでに鳥類が予想以上に多様化していたことが明らかになっている。飛行という適応は、新種の急速な進化を促進したのである。

鳥類の飛行はエネルギー消費が高い活動である。鳥類は比較的小型で内温性であり、飛ぶために多くのエネルギーを消費するため、大量の酸素が必要である。そのため、気嚢式の肺は鳥類にとって重要な助けとなっているのである。
恐竜の生殖と酸素濃度
20世紀の古生物学が誇る重要な成果の一つに、「恐竜の卵の発見」が挙げられる。20世紀後半に複数の卵化石が発掘されたとき、卵化石が示す複雑なパターンから、恐竜の生殖行動が単純ではないことが示唆された。特に産卵行動についてはその特性が認識されている。
現世紀に入り、新しい機器、すなわち小型の卓上スキャナーの利用が可能になり、恐竜の生殖に関する理解が大きく進展したと言える。今日、卵の内部の微細な胚を損傷することなく調査することが可能になったため、胚の成長と卵自体の構造の両方についての知識が深まっている。結果的に、恐竜の卵がどのように機能し、何故そうなったのかが解明されつつある。
現生鳥類は、少なくとも生殖に関しては恐竜と大きな違いが見られない。
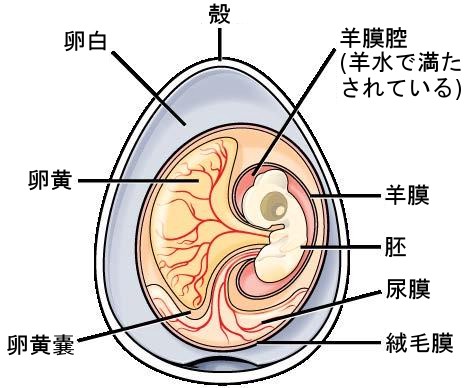
- 恐竜を理解するための理想的な指標である現生鳥類は、すべて炭酸カルシウムで構成された多孔質の卵を産む。
- これは、胎生を持つ種が多い現生爬虫類とは対照的である。
- また、鳥類と一部の爬虫類の卵の形状には顕著な違いが存在する。両者の卵殻は二重構造で、内側の有機膜が外側の結晶層に覆われているという共通点があるが、結晶質の量には大幅な差が存在する。鳥類の卵のように炭酸カルシウム層が厚いものもあれば、結晶質をほとんど含まないものも存在する。後者では外側の
層が膜状となり、革のように弾力性を持つ。
- さらに、結晶層の鉱物の種類も、鳥類・ワニ類・爬虫類の「方解石」から、カメ類の「アラレ石(炭酸カルシウムの別の結晶形)」まで多種多様である。
このように卵は、「硬質の結晶質卵」と「柔らかな羊皮紙状の卵」の二つの大きなタイプに分類される。
一部の研究者は羊皮紙タイプをさらに詳細に分類し、弾力性のあるもの(一部のカメ類とトカゲ類)と、柔らかなもの(大部分のヘビ類とトカゲ類)に区分している。化石化の可能性は、卵の硬さの度合いに大きく依存する。硬質の卵の化石(主に恐竜のもの)は大量に発見されており、弾力性のあるタイプの化石も一定数確認されているが、明らかに柔らかいタイプのものは全く残されていない。
恐竜に対する広範な興味から、その生殖習性についても多くの推測がなされてきたが、まだ未解明な点が多い。しかしながら、恐竜が卵生だったという確認は革新的な発見であった。恐竜の卵は大きく、方解石結晶(炭酸カルシウム成分)の層で覆われていた。これは、1920年代にアメリカ自然史博物館が初めてゴビ砂漠への探検旅行を行った際に発見された。以降、白亜紀の恐竜の卵は何千個も掘り出されており、モンタナ大学のジャック・ホーナーによる巣作りのパターンの発見と公表により、恐竜の生殖行動を推測することが可能となった。
しかし、これらの知見はあくまで白亜紀に関してのものであり、恐竜全体の特性と見なすことが適切であるかは疑問の余地がある。この未解決の問いは議論を呼んでいる。多くの研究者は、すべての恐竜が硬質の卵殻を持つ卵を産んだとの前提に立っているが、そのような証拠はまだ見つかっていない。これから見ていくとおり、初期の恐竜の一部は、羊皮紙タイプの卵を産んだか、あるいは胎生でさえあった可能性が、間接的ながら裏付けられている。
恐竜の卵の化石はほぼ全てが白亜紀の地層から産出している。卵の結晶形状、大きさや数、気孔のパターンなどの特性には大きなばらつきが見られる。しかし、科学的に最も興味深い謎はそのばらつきにあるわけではない。
- ジュラ紀の恐竜の卵が白亜紀のものよりはるかに少なく、三畳紀の卵はほとんど確認されていないという事実である。これは一体何故なのだろうか。
これまでに発見された恐竜の卵のほとんどは白亜紀のものである。前述の通り、これらは主に白亜紀後期の卵で、殻は炭酸カルシウムで構成されている。恐竜が卵を埋める習性も白亜紀後期の特性である。しかし、白亜紀よりも前の時代ではどのようだったのか。ジュラ紀後期の竜脚類や二足歩行の竜盤類の卵は存在する。特に優れているのはポルトガルで発掘されたもので、卵の内部には胚の骨も残っている。しかし、ジュラ紀後期よりも前の地層には、恐竜の卵や巣はほとんど見当たらない。三畳紀のものと確認された卵はご く少数である。
- したがって、どのような種類の卵がいつ初めて出現したのかは依然として不明確である。
- ペルム紀には無弓類(カメ類の祖先)、双弓類(ワニ類や恐竜の祖先)、さらに単弓類(我々ヒトの祖先)が生息していたにも関わらず、その時期の卵の化石は確認されていない。
- しかも、三畳紀後期の卵で、恐竜が産んだと思われるものはごくわずかしか知られていない。
- 私たちは大きな難問を突きつけられている。白亜紀の堆積物には恐竜の卵の化石が多数保存されているのに、ペルム紀や三畳紀の地層からは、同じような堆積環境であっても見つからないのである。主竜類(つまり当時の爬虫類)がペルム紀や三畳紀に硬い卵を産んでいたなら、まず間違いなくすでに発見されているはずである。
証拠の不在が不在の証明にはならないという点は警戒が必要である。それにもかかわらず、我々は結局、すべての証拠が示唆する事実を受け入れざるを得ない。
- 白亜紀より前の時代には、陸上動物が硬い卵を産むことは一般的ではなかったと、すべての証拠が示唆している。2012年に南アフリカで発見されたジュラ紀前期の恐竜の硬い卵は、その例外的な事例であった。
- これからどんなに熱心に化石収集が行われたとしても、現在の状況、すなわち硬い卵が見つからないという流れを覆す可能性は極めて低いと考えられる。
「理想の」酸素濃度
進化についての興味深い新発見として、
- 現代の陸上動物の祖先系統の多くが比較的短期間に現われたという事実が挙げられる。
- 具体的には、「酸素濃度が現在よりも高かった古生代後期(石炭紀~ペルム紀)」における現象である。
これは現生脊椎動物の多くの分類群、そして後のトカゲ類、カメ類、ワニ類、哺乳類につながる「科」の最初の動物にも当てはまる事実である。また、この傾向は陸生脊椎動物に限った話ではない。陸生無脊椎動物の多くも三億年以上前の石炭紀に出現している。これには多くの昆虫やクモ類、そして陸生の巻貝類(カタツムリやナメクジなど)の祖先系統も含まれている。過去5年間に行われた新たな実験により、
- 陸生脊椎動物の卵と昆虫の卵にとって、胚の発達が最速となる「理想的な酸素濃度」が存在することが確認された。
- その値は『27パーセント』である。
現在の大気中の酸素濃度は21パーセントであるが、ワニ類と昆虫に対する研究によれば、理想的な発達は27パーセントの酸素濃度時に示される。それ以上でもそれ以下でも、卵の孵化までの時間は長くなる。酸素濃度が10~12パーセントに低下すると、多くの卵が孵化しないか、孵化しても時間がかかりすぎて捕食される可能性が大幅に増える。ここに高温という要素が加わると、生存率はさらに下がる。なぜなら、卵には酸素を取り入れるための孔が必要であり、それが原因で水分が失われ、胚が死亡する可能性が高まるからである。
もし世界が現在よりも高温で乾燥しており、さらに酸素濃度が10~12パーセントであったなら、それは最悪の状況と言えるだろう。我々がそのような時代を認識しているのが三畳紀後期である。この時代、卵を産む動物は困難な状況に直面していたのである。
問題となるのは、爬虫類が出現した石炭紀が高酸素環境であったという事実である。その時の酸素濃度は27パーセントを超えていた。この初期の爬虫類は、他に先駆けて卵に羊膜を持っていた。ところが、地球規模で酸素濃度が下がって気温が上がると、その卵の構造が裏目に出た可能性がある。というのも、卵の外側から酸素が十分に浸透してこないにもかかわらず、内側からは過剰な水分が滲み出していたからである。
高温と低酸素環境(特に高温ではその傾向が強まる)に対応するためには、卵を産むよりも子供の形で直接産む方が適している可能性がある。
- すなわち、『胎生』が誕生したのは、ペルム紀後期に全球的に酸素濃度が低下していたことに対する適応だったと考えられる。
- 南アフリカ、ロシア、南米などで、多数の獣弓類の骨が発見されているにもかかわらず、同じ地層から卵や巣が見つかった記録はない。このことから、獣弓類はすでに胎生を進化させていた可能性がある。
- そして、この特性は獣弓類の子孫である真の哺乳類に受け継がれ、最初の哺乳類は最初の恐竜が現れた時期とほぼ同時に出現した。
恐竜の様々な系統がジュラ紀後期に炭酸カルシウムの卵を進化させたのは、酸素濃度が上昇したからである可能性がある。ペルム紀後期からジュラ紀中期までは大気中の酸素濃度が低かったため、炭酸カルシウムのような構造の卵を埋めてしまうと、中の胚は成育できなかったのである。
まとめると、
- ペルム紀後期から三畳紀にわたる低酸素で高温の環境は、胎生と柔らかい卵の進化を促した可能性が考えられる。柔らかい卵は酸素の取り込みや二酸化炭素の排出が容易であった。
- 一方、ジュラ紀後期から白亜紀にかけての高酸素で高温の環境に対応して、恐竜は硬い卵を進化させ、複雑な構造の巣の中に卵を埋めるようになったのである。
図表
図173 プレシオサウルスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「プレシオサウルス」(https://ja.wikipedia.org/wiki/%E3%83%97%E3%83%AC%E3%82%B7%E3%82%AA%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9)
図174 三畳紀・ジュラ紀・白亜紀における恐竜の属数の変化
A comprehensive resource for researchers in the Earth Sciences
(https://pubs.geoscienceworld.org/gsa/geology/article-abstract/32/10/877/29382/Shape-of-Mesozoic-dinosaur-richness?redirectedFrom=fulltext)
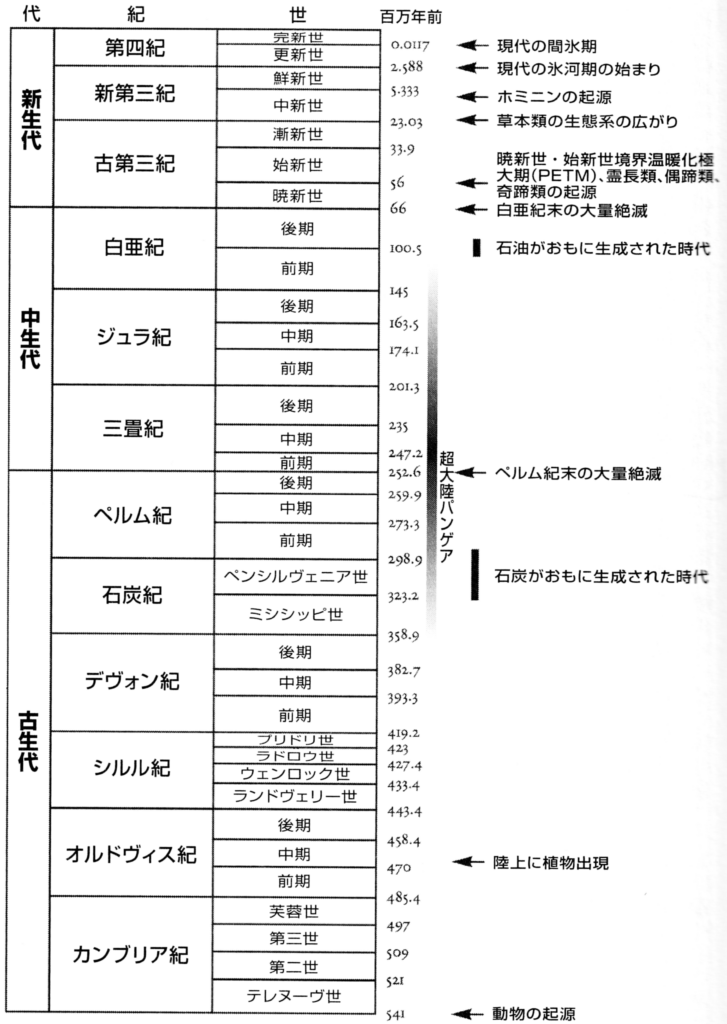
図175 地球史年表(中生代~新生代)
フリー百科事典『ウィキペディア(Wikipedia)』「カンパニアン」(https://ja.wikipedia.org/wiki/%E3%82%AB%E3%83%B3%E3%83%91%E3%83%8B%E3%82%A2%E3%83%B3)
図176 被子植物の構造
Qikeru(キケル)
(https://media.qikeru.me/angiosperms-gymnosperms/)
図177 スタウリコサウルス
Weblio 辞書「スタウリコサウルス・プリセイ」(https://www.weblio.jp/content/%E3%82%B9%E3%82%BF%E3%82%A6%E3%83%AA%E3%82%B3%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9%E3%83%BB%E3%83%97%E3%83%AA%E3%82%BB%E3%82%A4)
図178 ヘレラサウルス
恐竜ネット
(https://kyouryu.net/herrerasaurus/)
図179 二足歩行ワ二・バトラコプス・グランディスの想像図
ナショナル ジオグラフィック日本版
(https://natgeo.nikkeibp.co.jp/atcl/news/20/061500353/)
図180 ユーパルケリアの想像図
educalingo
(https://educalingo.com/ja/dic-ko/e-upaleukelia)
図181 爬虫類の展開
鳥取大学医学部保健学科検査技術科学専攻のブログ
「爬虫類の展開!」2015-03-05 20:05:15
(https://ameblo.jp/torii-hoken-kensa/entry-11997208537.html)
「鳥頸類」を加筆
図182 距骨の例(ドロマエオサウルス類)
福井県立恐竜博物館、恐竜博物館の調査研究、2008年7月29日
(https://www.dinosaur.pref.fukui.jp/research/200807excavation/)
図183 エウディモルフォドンの想像図
恐竜図鑑「エウディモルフォドン」
(https://kyouryu.info/eudimorphodon.php)
図184 ラゴスクスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「ラゴスクス」(https://ja.wikipedia.org/wiki/%E3%83%A9%E3%82%B4%E3%82%B9%E3%82%AF%E3%82%B9)
図185 恐竜の分類
福井県立恐竜博物館、恐竜・古生物 Q&A「恐竜の分けかたは?」
(https://www.dinosaur.pref.fukui.jp/dino/faq/r02003.html)
図186 アロサウルスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「アロサウルス」
(https://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%AD%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9)
図187 ハドロサウルスの想像図
恐竜のデジタル図鑑「ハドロサウルス」
(https://www.dd-lib.net/sort/kobetsu.cgi?sn=1233574859_963101137)
図188 イグアノドンの想像図
恐竜のデジタル図鑑「イグアノドン」
(https://kyouryu.info/iguanodon.php)
図189 角竜類・トリケラトプス
フリー百科事典『ウィキペディア(Wikipedia)』「トリケラトプス」
(https://ja.wikipedia.org/wiki/%E3%83%88%E3%83%AA%E3%82%B1%E3%83%A9%E3%83%88%E3%83%97%E3%82%B9)
図190 パンティドラコの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「パンティドラコ」
(https://ja.wikipedia.org/wiki/%E3%83%91%E3%83%B3%E3%83%86%E3%82%A3%E3%83%89%E3%83%A9%E3%82%B3)
図191 恐竜の基本的な3つの体型
新潟県立自然科学館、科学館日記、2017年5月4日NIIGATA-SCIENCE-MUSEUM投稿
「恐竜の分類が変わるかも!?」
(https://sciencemuseum.jp/cp-bin/wordpress/2017/05/04/)
図192 古生代以降の地質時代区分
「世界の起源 人類を決定づけた地球の歴史ORIGINS How the Earth Made Us」
ルイス・ダートネル Lewis Dartnell著、東郷えりか訳、株式会社河出書房新社、
2019年11月30日 初版発行
49頁「地球の地質時代区分」
図193 恐竜の系統図
コトバンク、日本大百科全書(ニッポニカ)「恐竜」
(https://kotobank.jp/word/%E6%81%90%E7%AB%9C-53115)
図194 コエルロサウルスの想像図
iStockのデザイン「コエルロサウルス類」
(https://www.istockphoto.com/jp/search/2/image?phrase=%E3%82%B3%E3%82%A8%E3%83%AB%E3%83%AD%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9%E9%A1%9E)
図195 ステゴサウルスの想像図
恐竜図鑑「ステゴサウルス」
(https://kyouryu.info/stegosaurus.php)
図196 トリケラトプスの想像図
恐竜図鑑「トリケラトプス」
(https://kyouryu.info/triceratops.php)
図197 ティラノサウルスの想像図
恐竜図鑑「ティラノサウルス」
(https://kyouryu.info/tyrannosaurus.php)
図198 パンゲア超大陸とその分裂
コトバンク「パンゲア」・デジタル大辞泉「パンゲア」の意味・読み・例文・類語
(https://kotobank.jp/word/%E3%83%91%E3%83%B3%E3%82%B2%E3%82%A2-606287)
図199 裸子植物(左)と被子植物(右)の胚珠
Qikeru(キケル)
(https://media.qikeru.me/angiosperms-gymnosperms/)
図200 恐竜と鳥類の系統樹
川崎悟司@satoshikawasaki
(https://twitter.com/satoshikawasaki/status/1421722216253706245)
図201 トロオドンの想像図
丹波地域恐竜化石フィールドミュージアム「トロオドン類」
(https://tamba-fieldmuseum.com/study/fossil/troodon)
図202 ドロマエオサウルスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「ドロマエオサウルス」
(https://ja.wikipedia.org/wiki/%E3%83%89%E3%83%AD%E3%83%9E%E3%82%A8%E3%82%AA%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9#:~:text=%E3%83%89%E3%83%AD%E3%83%9E%E3%82%A8%E3%82%AA%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9%EF%BC%88Dromaeosaurus%EF%BC%89%E3%81%AF,%E3%83%88%E3%82%AB%E3%82%B2%E3%80%8D%E3%81%AE%E6%84%8F%E5%91%B3%E3%81%A7%E3%81%82%E3%82%8B%E3%80%82)
図203 始祖鳥の想像図
フリー百科事典『ウィキペディア(Wikipedia)』「始祖鳥」
(https://ja.wikipedia.org/wiki/%E5%A7%8B%E7%A5%96%E9%B3%A5)
図204 プテロダクティルスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「プテロダクティルス」
(https://ja.wikipedia.org/wiki/%E3%83%97%E3%83%86%E3%83%AD%E3%83%80%E3%82%AF%E3%83%86%E3%82%A3%E3%83%AB%E3%82%B9)
図205 エオアルラヴィスの想像図
Wikipedia, the free encyclopedia“Eoalulavis”
(https://en.wikipedia.org/wiki/Eoalulavis)
図206 有羊膜類の卵の構造
生物学 第2版 — 第29章 脊椎動物 —,OpenStax, Rice University,6100 Main St., Houston,
TX 77005-1827にて公開されている教科書“ Biology 2e”の日本語訳
(https://medium.com/@BetterLateThanNever/%E7%94%9F%E7%89%A9%E5%AD%A6-%E7%AC%AC2%E7%89%88-%E7%AC%AC29%E7%AB%A0-%E8%84%8A%E6%A4%8E%E5%8B%95%E7%89%A9-bb7412be3b72)
