
第四部 生命の進化
第8章 大絶滅と三畳紀爆発【2億5200万 ~ 2億年前】
1.大絶滅 ― 酸素欠乏と硫化水素【2億5200万 ~ 2億5000万年前】
南アフリカ中部に位置するカルー砂漠には、約2億7000万年前から約1億7500万年前にかけて堆積した岩石が見受けられる。これらの岩石群の中から、保存状態の良好な大型陸上動物の化石が発見された。大量絶滅の中でも最も影響の大きかった『ペルム紀-三畳紀(P-T)境界絶滅』の前後に生きていた動物群である。
厚さ千数百メートルにもなるカルーの堆積層を上っていくと、時間の進行とともに気温や酸素濃度の変動が見受けられる。始点は氷原や氷山が主流だった時代で、終わりには地球史上極めて暑いと推測される時代に至る。さらにその過程において、約6億年前の動物の誕生以来、大気中の酸素濃度が最も低下した期間が数千万年間にわたり存在する。この全体像から多くの学びが得られるが、中でも特に詳細に研究されている時期が存在する。
それは2億5200万年前から2億4800万年前の岩石層で、厚さは数百メートルに及ぶ。ペルム紀末期の1000年と、それに続く2億5200万年前の大規模な大量絶滅の最初の数百万年間に形成されたものである。
この地層からは、数は限られるが非常に良好な保存状態の頭骨や体骨格が見つかる。地球科学者たちは数十年前から、これらの化石や岩石に対して数多くの重要な問いを投げかけてきた。
- 一つ目の問いは、大量絶滅がどれほどの期間にわたり続いたのか
である。起点となるのは、絶滅率が通常の「自然な」絶滅率(およそ5年に一回と推定されている)を最初に上回った時点である。
- 二つ目の問いは、ペルム紀においては陸上と海洋で同時に大量絶滅が起こったのか
ということ。
- 三つ目の問いは、おそらく最も興味深い謎だが、大量絶滅の原因は何であったのか
という点である。そして、
- 最後の問いは、陸上の生態系がどれだけ速く回復したのか
という問題であり、これを突き止めることには大きな意味がある。それがわかれば、人類がいずれペルム紀級の大量絶滅に見舞われても生き延びるための手がかりが得られるかもしれないからである。そういう未来が訪れる可能性は、我々が考えているよりも高い可能性がある。
ペルム紀絶滅の余波
『ペルム紀の大量絶滅』をめぐっては、その原因について今なお激しい論争が繰り広げられている。しかし、一つの点に関しては意見の一致を見ている。
- 絶滅後の生態系が深刻なダメージを受け、その回復が大幅に遅れたという事実である。後の『白亜紀-第三紀(K-T)境界の絶滅』と比べて、ペルム紀の場合はこの点が大きく異なる。どちらも地球上の種の半数以上を消滅させたにもかかわらず、「K-Tイベント」の後は世界が比較的速く回復した。
これは二つの絶滅の原因が異なることが理由であるかもしれない。
- 「K-Tイベント」の場合、小惑星の衝突とそれによる環境破壊が原因であり、これは10年以上前から認められている。しかし、この衝突による混乱状況はすぐに収束した。
- ペルム紀絶滅の場合は異なる。先に述べた通り、ペルム紀の絶滅を引き起こしたのも大型隕石の衝突だと主張する地球科学者も存在するが、K-T境界と異なり、絶滅を引き起こした環境条件は絶滅開始から数百万年にわたり続いたとされる。回復の兆しが見られるのは、約2億4500万年前の三畳紀中期になってからである。
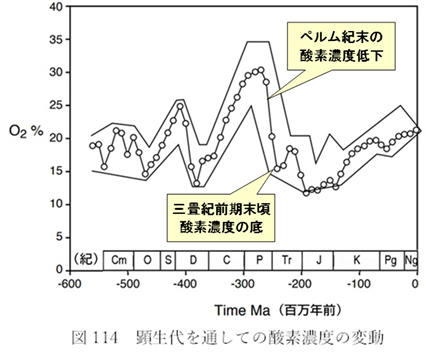
これらの結果から、ペルム紀末の酸素濃度の低下が、直接的または間接的に大量絶滅の一因となったと考えられる。最新の『バーナー曲線」』(<物語り その10>図116)によれば、酸素濃度は三畳紀に入っても低いままで、上昇に転じるのは三畳紀前期の終わり頃であった。これが回復が大幅に遅れた理由の一つであると解釈できる。
これらのデータから推測できるのは、絶滅を引き起こした環境的事象が持続していたということである。もしこれが事実であり、さらにはそのような有害な状況に適応する能力が動物にあったとすれば、
- 三畳紀には多数の新種が出現したと予想される。
- 大量絶滅によって空になった生態系のニッチを埋めるためだけでなく、絶滅自体の影響が長引いたことによる環境への反応として新種が出現した可能性もあるからである。
実際、三畳紀についてはこのようなパターンが確認されている。多くの新種が世界に補充されたのである。
- その中には、絶滅した種と外見や行動が似たもの(すなわち、同じニッチを占める種が入れ替わった状況)も少なくない反面、
- とくに陸上ではまったく新しい生物も多数誕生した。
次章ではこの後者の種に焦点を当て、その多くが長期間の低酸素状況に対応するために進化したということを説明する。この低酸素期は三畳紀の終わり頃に始まり、ジュラ紀に入ってからも続き、5000万年あまりの長きにわたったのである。
- 三畳紀は、二つの異なる環境に適応した動物たちが交差した時代と言える。一つは酸素濃度の高い環境、もう一つは酸素濃度の低い環境であった。
ペルム紀絶滅の原因 - 隕石衝突か温室効果か
20世紀が終わり21世紀が始まると、ペルム紀絶滅への注目度はこれまでにない高さに達した。
その理由は、これまでの大量絶滅の中で最も被害が甚大だったという事実による。実際に、全種の90%が絶滅したと推定されており、この数値は現在、あらゆる文脈で頻繁に引用されているのである。
しかし、絶滅の原因を究明するには、どれほど多くの種が絶滅したかよりも、どれほどの期間で絶滅が起きたかを明らかにすることが重要である。これに関して大いに参考になる化石が、中国とアメリカの古生物学者による大規模な研究調査によって明らかになった。調査の対象となったのは、ペルム紀から三畳紀にあたる厚い石灰岩で、中国南東部のメイシャン(煤山)近くに露出していたのである。
ペルム紀絶滅の原因については、これまで数多くの可能性が提唱されてきたが、2005年にペンシルベニア州立大学の地球化学者チームから新たな解答のヒントが提供された。この研究チームを率いるリー・カンプは、海洋の化学についての世界的な専門家であり、特に海洋の炭素循環に関する権威である。カンプは、長年の同僚であるマイケル・アーサーと共に論文(注141)を発表し、
- ペルム紀末の海に存在した微生物(具体的には紅色硫黄細菌とは異なる種)が生成した『硫化水素』が、陸地と海洋の両方での絶滅に直接関与した可能性を提唱した。
(注141)Lee R.Kump and Michael A.Arthur,” Interpreting carbon-isotope excursions: carbonates and organic matter”, Chemical Geology 161(1999) 181-198
カンプの仮説 - 温室効果絶滅説の始まり
カンプらが提唱するシナリオは次の通りである。
通常の海洋環境では、現代の黒海のように酸素濃度の高い表層水と硫化物濃度の高い深層水は分離している。しかし、海洋が低酸素状態にあった時期(特に海底だけでなく海面近くでも酸素が不足していたとされる時期)には、深海の硫化水素濃度が一定の限界を超えると、その有毒な深層水が急激に海面に浮上することがあり得る。その結果、大量の有毒な硫化水素ガスが大気中に放出される可能性がある(注142)。
(注142)より丁寧な説明を行うと、まず、大規模な火山活動によって膨大な二酸化炭素とメタンが放出され、急激な地球温暖化を引き起こす。そうすると、海水温が上がって大気から海に溶け込む酸素の量が減る。これに伴い、深海の「硫酸塩還元細菌」(注143)が作り出した硫化水素に満ちた水が表層に湧き上がり、硫黄細菌が繁殖するとともに酸素呼吸をする海生生物が窒息死する。硫化水素は大気にも拡散し、陸上の動植物の命を奪うほか、[H2S+O3 → S+H2O+O2]なる反応によって上空のオゾン層を破壊する。オゾン層の保護が失われ、太陽の紫外線によって残りの生命も死に絶える。
(注143)「硫酸塩還元細菌」は嫌気的環境で有機物を分解し、そこで生じた電子を用いて硫酸塩を還元する微生物の総称であり、嫌気的環境に広く分布し、硫黄の循環に重要な役割を担っている。したがって、硫酸塩還元細菌の餌は「硫酸塩」で、硫酸塩を硫化物イオンにまで還元し、硫化水素を発生したり、金属イオンとの硫化物を生成する。
地球規模の絶滅の原因が「硫化水素」であったとすると、海洋と陸上の両方で絶滅が起こったことも理解できる。たとえ海洋から大気への硫化水素の放出が比較的穏やかだったとしても、対流圏に一定量蓄積すれば、それは動植物にとって致命的な濃度に達する可能性があるからである。このシナリオのようなことはペルム紀末だけでなく、地球史におけるほかの時期でも起きた可能性があり、大量絶滅を引き起こす重要な要因の一つだったのかもしれない。
カンプらの研究チームが行った大まかな計算によれば、
- 驚くべきことに、ぺルム紀後半の大気に入り込んだ硫化水素ガスの量は、現代の小規模な流入量(火山由来の有毒ガス)の2000倍以上であった。大量の硫化水素が大気中に放出され、有毒なレベルに達したことは、間違いなく推定される。
- さらに、生命を危険なレベルの紫外線から守る「オゾン層も破壊された」と推測される。この推測には証拠が残されており、絶滅の時期にあたるグリーンランドの堆積層から取得した胞子の化石には、突然変異の形跡が認められた。オゾン層の喪失とその結果、生物が大量の紫外線に長期間晒されたと考えると、このような結果は不思議ではない。
さらにもう一つ、メタンガスの濃度が急激に高まって100ppmを超えると、それに伴う二酸化炭素の蓄積と相まって地球温暖化が著しく増幅される。
硫化水素は大気中に流入しながらオゾン層を破壊し、同時に温室効果ガスが地球を温める。硫化水素の致死性は、気温の上昇とともに増すことが確認されている。これらの研究結果により、隕石衝突説に代わる新説が提起され、その信頼性は高そうに思われる。このようなメカニズムに基づいているとすれば、絶滅は長引いたとも推測できる。あるいは短い絶滅イベントが断続的に起きて、そのたびに生物を死滅させていったとも考えられる。
これまでの視点は、岩石自体から得られる証拠に焦点を当ててきた。しかし、過去の出来事を解明するにはもう一つの方法が存在する。それは過去の「大気組成をモデル化」する方法である。ペルム紀に関しては、酸素と二酸化炭素の濃度に加え、当時の大気温度のモデルも作成されてきた。イェール大学のロバート・バーナーは大気中の二酸化炭素濃度と酸素濃度の変遷を計算し、ペルム紀に酸素濃度が急落し、それに伴って二酸化炭素濃度が著しく上昇したことを確認した。
次にカンプらのグループが困難な研究に挑み、地球全体で硫化水素が放出されそうな場所の分布を明らかにすることを試みた。そのために彼らは、『大気大循環モデル(GCM)』(注144)を利用した。
(注144)『全球気候モデル(Global Climate Model, GCM)』は、気候モデルの一つで、大気・海洋・陸地・雪氷などの変化を考慮し、流体力学・力学・化学・物理学・生物学などの方程式を用いて地球の気候を再現し、気候の変化を表現する数理モデルの総称である。主に大気循環、つまり大まかな気象現象を再現することを目的としている。『大気循環モデル(General Circulation Model, GCM)』、『大気大循環モデル』とも呼ばれる。
この種のモデルは、もともと現代の天候や気候のパターンを理解するために開発されたものである。しかし、ペルム紀末から三畳紀にかけての時期に関しては、各大陸の位置が判明しており、大気中や海中の酸素濃度や二酸化炭素濃度、さらに大気温度や海水温度もわかっているため、GCMはペルム紀にも適用可能である。
- 硫化水素の発生は一度きりではなく、繰り返し起こり、特に世界中でP-T境界層が堆積していた時期に集中していた。
- カンプのモデルは、硫化水素が海から大気中に湧き上がる場所だけでなく、最終的にどれだけの硫化水素ガスが大気に入り込んだかを計算し、初期の試算を裏付けた。その結果、ほとんどの陸上生物を死滅させるに十分すぎる量の硫化水素が大気に流入したと結論付けられた。
- 加えて、この有害なガスは海水にも溶け、浅海の環境に生息する生物に対して致命的な影響を及ぼした。特に、サンゴ、二枚貝、腕足動物、外肛動物などの炭酸カルシウムの骨格を分泌する無脊椎動物は甚大な打撃を受けたと考えられる。
標高圧縮
『標高圧縮』という現象について考察すると、酸素濃度の変動についても、かつては未認識だった一面が明らかとなっていることが判明する。
この変動は種の移動や遺伝子流動に影響を与える。山脈は遺伝子交換の妨げとなりやすく、その結果、山脈の両側には異なる生物相が形成される。ペルム紀末には酸素濃度が低下していたため、海抜ゼロメートルの地点でも、まるで標高5000メートルで呼吸するかのような状況であったと推測される。このため、ペルム紀の期間中には、標高が高くなくても呼吸が困難となり、山脈どころか丘陵程度の地形でも生物の往来が制限された。その結果として、海抜ゼロメートルの海岸線に沿って、固有種が発生する地域が点々と連なる世界が広がっていたと推察される。
高地に適応する種を除けば、多くの大陸の高原では生物が生息していなかった可能性がある。ペルム紀末の大陸の位置を考慮すると、これは予想外の事態である。2億5000万年前にはすべての大陸が結合し、一つの巨大なパンゲア超大陸となっていた(192頁、図142参照)。動物は大西洋に阻まれることなく超大陸を横断できたはずだから、生物相の特徴を基にした地理的区分、つまり生物地理区の数はごくわずかだったと推察される。しかし、実際には、
標高が種の移動の新たな障壁となった。様々な脊椎動物相に関する最新の研究を考慮すると、陸上はいくつもの生物地理区に分かれていたと推定される。
20世紀末から21世紀初頭にかけて、ペルム紀末に関する調査がいくつか実施された。ピーター・ウォード、ロジャー・スミス、ジェニファー・ボータの研究チームはカルー砂漠で調査を行い、マイケル・ベントンはロシアで、さらにクリスチャン・シドーはニジェールで調査を行った。その結果、
アフリカでは地域によって動物相が明確に異なり、ほとんど重複しないことが確認された。つまり、低酸素状態の時期には、標高が大きな障壁となり、種の移動や遺伝子流動を阻害したのである。したがって、少なくとも低酸素期の陸上はいくつもの生物地理区に分かれていたと考えられる。一方、酸素濃度の高い時期には逆の現象が起きた。すなわち、生物地理区の数は少なく、世界中に広く分布する動物相も存在したと推定される。
- 酸素濃度の急落は、種の移動を妨げるという意味では新山脈ができる以上の効果を持っていた。ペルム紀後期から三畳紀を通して、標高1000メートルを超える地域の大半で生物が棲めなくなったのである。この効果は『標高圧縮』と呼ばれ、酸素濃度が底を打った時代には三畳紀の陸上動物に大きな打撃を与えた可能性がある。
「標高圧縮」によって生息地が奪われた場合、動物は高地から海抜ゼロメートルの地域への移動を余儀なくされ、それが不可能な場合は絶滅したと考えられる。また、移動に伴って、空間や資源を巡る競争が激化した。新たな捕食者、寄生虫、病気が生物相の豊かな低地に侵入し、それによって多くの種が絶滅した可能性がある。
- ピーター・ウォードらの推定によれば、ペルム紀末には標高圧縮の影響で地表の半分以上が生息不能となっていたと考えられる。
何十年も前、ロバート・マッカーサーとE・O・ウィルソンは著書『島の生物地理学説』で一つの絶滅モデルを提唱した。彼らが指摘したのは、
- 種の多様性は生息域の面積に関係し、島や保護区などの面積が小さいほど、種の絶滅が起きやすいという事実である。
- だとすれば、ペルム紀末にはそれと同じ効果で絶滅するケースがあったかもしれない。標高圧縮が起きれば、結果的に大陸塊上の利用可能域が狭まるからである。
ペルム紀絶滅の惨状
最後に、ペルム紀絶滅についてもう一つの特徴を取り上げる。それこそがペルム紀大量絶滅というテーマの核心に迫るものだからである。
ピーター・ウォードとその大学院生、フレデリック・ドゥーリーは、リー・カンプと共同研究を進め、驚くべき発見をした。ドゥーリーは硫化水素が植物や一部の動物に与える影響を調査しており、カンプは一方でペルム紀末の海洋状況のモデル化を行い、地球全体の海洋表層水に存在する硫化水素の量も算出していた。カンプの結論に基づき、ドゥーリーは試験を実施した。試験対象は単細胞の海洋性植物プランクトンと、海洋性動物プランクトンとして重要な役割を果たすカイアシ類(エビに似た微小生物)であった。実験で用いた硫化水素の濃度では植物プランクトンは死滅せず、意外にも逆にその成長が加速した。しかし、カイアシ類はほとんど瞬時に死滅した。通常、カイアシ類が植物プランクトンを餌にして、その数を抑えるのだが、この場合、カイアシ類がいないので、
- 植物プランクトンは海底に沈み、腐敗して最後の酸素までを奪い取った。
- このような状況が生じれば、炭素同位体のパターンの大きな変動はもちろん、幼生の一時期にプランクトンとして海面近くで生活する全ての海洋動物種が消滅するであろう。
- 最終的に地球は腐敗した植物で窒息状態となり、ほとんどの動物が存在しなくなる。これがまさにペルム紀末に起こったと考えられる。少なくとも海洋ではそうであった。
また、P-T境界期における陸上動物の絶滅に関しては、著者らがカルーで行った脊椎動物の研究(2005年に発表)が現時点では最良の記録といえる(注145)。一方、南アフリカのロジャー・スミスの研究により、2億5200万年前の南アフリカでは異常な乾燥と急激な気温上昇が起きていたことが明らかになった。これは、非常に信頼性の高い証拠に基づいている。スミスは、干ばつと高温だけでも、大半の脊椎動物が絶滅した理由となり得ると主張している。大気中と海中の有毒な硫化水素により、砂漠に死が訪れたのである。
(注145)P. Ward et al., “Abrupt and Gradual Extinction Among Late Permian Land Verrebrates(陸上脊椎動物) in the Karoo Basin(盆地), South Africa,” Science 307 (2005):709-714.
2.三畳紀爆発 - 2億5200万 ~ 2億年前
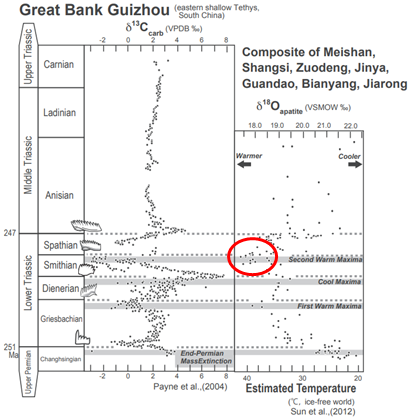
2012年、中国とアメリカの共同研究チームが驚くべき論文(注146)を発表した。それは、ペルム紀絶滅後の海洋生物の回復が大幅に遅れた理由を探究するもので、
- 当時の海水温度が40℃、陸上では60℃という焼けつくような暑さであったという結論を導き出していた。
(注146)YADONG SUN, MICHAEL M. JOACHIMSKI, PAUL B. WIGNALL, CHUNBO YAN, YANLONG CHEN, HAISHUI JIANG, LINA WANG and XULONG LAI, “Lethally(致死的に)Hot Temperatures During the Early Triassic Greenhouse,” Science 338 (2012):366-370.
この研究では、15,000点を超える標本を分析しており、ペルム紀絶滅後の環境条件に関して、これほど詳細で徹底した調査がなされたことは過去になかった。
この共同研究チームは、太古の高温の世界がどのようなものであったかを推測した。彼らが突き止めたような水温では、ほとんどの海洋生物が死滅したであろう。そのような環境下では、熱帯地方には動物がほとんど存在しなかったと推測され、複雑な生物は高緯度地方でのみ、僅かに生存していたと推測される。陸上動物は中緯度地方でさえも稀であったと考えられる。その暑さから、大気中には大量の水蒸気が存在し、熱帯地方では年間を通じて雨が多かったと推定される。しかし、雨が降ったとしても、植物が全く生えていなかったため、その不毛さから砂漠と変わらなかったと考えられる。
地質年代学は絶えず進展を遂げており、現在では、
- この高温期が『三畳紀』の開始から300万年以上続いたことが判明している。
- さらに、この期間中に温度が一段と上昇し、『スミシアン期』と称される時期(約2億4700万年前の100万年間)にはピークを迎えた可能性がある。これは動物が出現して以降、既知の中で最も高い温度であった。
ペルム紀絶滅が、全ての事象の中で最も壊滅的であったことは間違いない。しかし、それは多細胞の動物から見た場合の話である。微生物、特に硫黄を好み、酸素を嫌う微生物にとっては、この絶滅はまさに楽園が戻ってきたかのようであった。なぜなら、動物が出現するまでの間、全生命の大多数を占めていたのはその種の微生物であったからだ。
私たちは遠い未来から俯瞰しているので、
- ペルム紀絶滅はデボン紀末の出来事の再現であったと理解できる。どちらもいわゆる『温室効果絶滅』であった。
- 同じタイプの絶滅が三畳紀末にはさらに何度も発生し、ジュラ紀や白亜紀にも複数回起きて、最終的には暁新世末(約6000万年前)における最後の温室効果絶滅として終息した。
- しかし、ペルム紀の事象ほど大規模なものはなく、また、絶滅後にこれほど多様な動物が世に生み出されたこともない。
ペルム紀絶滅後、世界は新しい生物の誕生で満ちていた。しかし、私たちに直接関わるのは二つの全く新しい系統で、それぞれ三畳紀終焉の頃までには既に繁栄と進化を遂げていた。
- 一つは『哺乳類』
- もう一つは哺乳類の長年の競争相手である『恐竜』である。
哺乳類を活性化し、恐竜への基盤を構築する上で、ペルム紀絶滅が果たした役割は小さなものではない。しかしながら、
- 三畳紀の恐竜と哺乳類は確かに極めて重要な陸上生物であったが、両者とも三畳紀爆発の後半に出現した。
- その結果、三畳紀を通じて体が比較的小さいまま(特に哺乳類は最大でもネズミ程度の大きさ)で、個体数と種の多様性も限定的だった。
- 恐竜の時代が始まるのは次の『ジュラ紀』からで、そして今も続く哺乳類の時代が本格的に始まるのは『新生代』を待たねばならない。
恐竜と哺乳類が登場する遥か以前には、非常に興味深い動植物が三畳紀の進化の舞台に立っていた。既に長期間存在している分類群の新しい型もあり、全く新参の生物もいた。古生代を生き抜いた生物とは根本的に異なる新しいデザインを持つものだ。
そんな風に混在していることから、
- 『三畳紀はまさしく時の交差点』と呼ぶにふさわしい。
- いくつかの点ではカンブリア爆発と共通点を持つ。最初の動物であるエディアカラ生物群が絶滅した後、新しい動物が様々な形を進化させ、カンブリアの海を満たすように、「ペルム紀絶滅後に生まれた空白を、多数の新しい生物が埋め尽くした」のだ。
- そして大カンブリア爆発の時と同じように、新奇な体制の多くは短期的な実験に終わり、より優れたデザインを持つ生物との競争や捕食により絶滅に追い込まれた。
- これほど多様な生物が新たに出現した時代は、『カンブリア紀』と『三畳紀』をおいてほかにない。
そこには二つの主要な要素が働いていたと推測される。
- 一つは、生物が全く住まない空間がペルム紀絶滅によって大幅に増加し、少なくとも一時的にはほとんどどんな新デザインでもうまくいったことである。
しかし、それと同等か、それ以上に重要な新たな視点がもう一つ存在する。
ペルム紀絶滅は全ての絶滅イベント中で最も破壊的だったため、絶滅直後の三畳紀の世界にはごく少数の生物しか残っていなかった。
- さらに、どの大気モデルを見ても、「三畳紀という長い時代を通じて酸素濃度が現代より低かった」ことが示されている。以前に見たように、低酸素状態、特に大量絶滅後の低酸素状態は、生物の異質性を増大させる。つまり、新しい形態の多様性が増加する
これら二つの要素が相まって、カンブリア紀以来最多となる新しい形態が生まれた。
この観点から、三畳紀はまさしく重要なカンブリア紀に匹敵すると認識されており、生物が急速に多様化したこの時期を『三畳紀爆発』と呼んでいる。
三畳紀前期と正式に称される期間は、約2億5000万年前から約2億4500万年前である。この時期、大量絶滅後の回復の痕跡はほとんど見られなかった。酸素濃度は驚異的な変動を経て、10~15パーセントという最低限のレベルまで低下した後、約2億4500万年前から約2億4000万年前までの少なくとも500万年間その状態が続いた。この期間には、炭素の同位体パターンに大きな変動が存在したという記録が残されており、これは炭素循環そのものが乱れていたことを示唆している。メタンガスが海洋や大気に放出されたり、小規模な絶滅が立て続けに起こったりしたことが推測されている。この点もカンブリア紀前期と非常に類似している。
これらの証拠から見えてくるのは、動物にとって極めて厳しい環境であった。硫黄細菌などの微生物は繁栄した可能性があるが、動物は長期間逆境を耐えなければならなかった。しかしながら、この困難な状況が進化や新たな展開を促す主要な要素となった。実際に、
この「酸素不足の状況から新種の動物が生まれ」、
- そのほとんどが酸素危機に対する高い適応能力を持つ呼吸器系を有していた。
- 陸上では、厳しい環境の中から二つの新しいグループ、すなわち『哺乳類』と『恐竜』が登場した。前者は獣弓類に取って代わり、後者は世界を支配することとなった。
三畳紀には陸上でも海でも多様性が顕著であった。海では『二枚貝』の新たな祖先系統が誕生し、絶滅した多くの腕足動物(注147)に取って代わった。
(注147)触手動物腕足綱の総称。軟体部は2枚の殻で包まれるが、殻は二枚貝類と違って背腹に位置する。多くは海生で他物に着生するか、砂泥中にもぐって生息。移動する能力はない。カンブリア紀初期にすでに500種以上存在し古生代に最も栄え、中生代以降は衰退したが、その後のあらゆる地質時代から知られ、シャミセンガイ(汽水域にすむ)、ホオズキガイ、チョウチンガイなど約70属250種が現存する。
一方、『アンモナイト』(注148)や『オウムガイ』の大幅な多様化により、活動的な捕食者が再び海洋を満たした。過去に生きた全てのアンモナイトのうち四分の一以上が三畳紀の岩石から発見されているが、三畳紀の期間はアンモナイト類が地球に存在した全時間のわずか一割に過ぎない。しかしながら、三畳紀のアンモナイトは形状や模様が古生代の祖先と全く異なっていた。その理由は、この種の動物が無脊椎動物の中でも特に低酸素環境への適応力が高かったからである。新種のサンゴである石サンゴが礁を形成し始め、多数の陸生爬虫類が海に戻った。

(注148)アンモナイト亜網(ゴニアタイト目・セラタイト目・アンモナイト目と進化していくアンモナイト全体を指す)はデボン紀はじめに出現し、その祖先はバクトリテス類(オウムガイ類のオルソセラス目からシルル紀に派生した種類)から進化したと考えられている。初期のアンモナイト類は、まっすぐな殻を持っていた。ゴニアタイト目は、デボン紀後期からぺルム紀まで繁栄したが、ぺルム紀末期までには絶滅してしまった。ぺルム紀中期に出現したセラタイト目は、ぺルム紀末期の生物の大絶滅を乗り越え、三畳紀を通じて繁栄した。しかし、三畳紀中期にはアンモナイト目も出現し、ジュラ紀、白亜紀を通して大繁栄することになるが、セラタイト目はそれに押されるように、三畳紀末には絶滅してしまった。また、隆盛を極めたアンモナイト目も白亜紀には減少しはじめ、異常巻き(不規則な形)や複雑に巻いたもの、鉤状のもの、一部巻いたものなど、様々な形が発達した。そして白亜紀の終わり(6500万年前)にはついに恐竜とともに完全に地球上からいなくなり絶滅してしまった。
しかし、新旧の体制が置き換わったり、多様な体制の実験が行われたりといった意味で、最も大規模な変化が起きたのは陸上であった。陸上でこれほど多種多様な動物が生息した時期は他にない。ペルム紀によく見られたものも存在した。

ペルム紀絶滅を生き延びた『獣弓類』が多様化し、三畳紀の初めに陸上の覇権をめぐって『主竜類』(現生ではワニ、鳥類、過去においては恐竜に代表される爬虫類の分類群)と競い合った。しかし、獣弓類の優位は長くは続かなかった。陸地を支配しようとする多種多様な爬虫類が獣弓類と闘い、また爬虫類同士でも争っていたからである。哺乳類型爬虫類からトカゲまで、そして「最初期の哺乳類」から「本格的な哺乳類」まで、三畳紀はまさに『動物デザインの壮大な実験場』だった。
表面的な観察からでは、哺乳類が純粋な爬虫類に対して競争で優位に立つことは理解できることであった。
- 主に、この時期の哺乳類は大部分が恒温性で、おそらく卵生であった恐竜と比べて顕著な子育て能力を有していたと考えられる。現在でも、恒温性の動物はその特性を維持しているのである。
- さらに、哺乳類は歯の形状を変化させることで、小さな種子から草、さまざまな種類の肉まで、多様な食物を摂取する能力を獲得した。
これは、結果として哺乳類が地球を支配する大きな要因の一つとなったである。
- それにもかかわらず、哺乳類は当時の競争には勝つことができなかった。哺乳類型爬虫類の絶滅を持って第一の哺乳類時代は幕を閉じ、次の章へと進むこととなった。次のステージに登場した哺乳類は、一世を風靡した先輩たちとは異なる特性を持っていた。
ケンブリッジ大学の脊椎動物の古生物学者、ローランド・スーキアスらはさらなる研究を進め、
ペルム紀大量絶滅後の空白期から三畳紀前期にかけて主に二つの生体系が出現したことを明らかにした。
- それらは四肢を使用する『四肢動物』と、二本の足しか使用しない『二足動物』である。
- さらに、約5000万年にわたる三畳紀からジュラ紀へと移行する過程で、「トカゲ類」が「哺乳類型爬虫類」よりも多くの種と形態で多様化したことも明らかにされた。多様性の指標である体のサイズも確実に増加し、その事実が初めて数値で裏付けられたのである。
スーキアスらの研究では、トカゲ類の成長が他のグループよりも速く、成熟するまでの期間が短いことも確認された。この「生殖までの時間」に差があることが、生物の繁栄に影響を及ぼした可能性がある。成長して生殖可能になるまでの時間が短ければ、大型の植物食動物や大型の捕食者といった生態的な役割への短期間の適応が可能となる。この時点で獣弓類はまだ十分な成長を遂げておらず、この競争には勝つことは難しかった。
三畳紀後期には、恐竜が生態系に定着し、ジュラ紀の大きさにまで成長し、広範囲に存在していたと広く誤解されているが、シカゴ大学の古生物学者ポール・セレノの研究によれば、これらの前提は間違っていたようだ。
恐竜の出現(約2億2100万年前)から三畳紀末(約2億100万年前)までの約2000万年間、恐竜と獣弓類の数は少なく、体の大きさも小さかった。獣弓類よりも恐竜の数が多かった可能性はあるが、全体的にどちらのグループも大いに繁栄していたとは言えない。
そのため、陸生の四肢動物全体が成功していなかったことから、海への適応がより有利であったと推測されている。実際、地球の歴史において、三畳紀ほど多くの陸生四肢動物が海へと進出した時代はないである。
動物の生体系は強い選択圧によって進化し改良されてきた。特に、「低酸素環境」で充分な酸素を取り込みつつ、食物の摂取、繁殖、競争を維持しなければならないという状況は強い圧力となった。
- ペルム紀の高酸素環境で高度な活動を行っていた生物が、三畳紀の低酸素期という厳しい状況下でも同等の活動を続けられるためには、十分な酸素供給が必要であった。
- 大気中の酸素の三分の二が失われたという、「きわめて切迫した選択圧」に直面した際、進化という爆弾の導火線に火が点いたのは間違いなく、それが三畳紀になって爆発した。三畳紀における体制の多様化が、カンブリア爆発による海洋動物の体制の多様化に似ているというのはその点にある。
- 前述の通り、カンブリア爆発はエディアカラ動物群の大量絶滅後に発生し、しかも当時の酸素濃度は現在よりも低かった。「新たな生物形態はこの低酸素環境によって引き起こされた」と考えられているである。
三畳紀における生物の復活

ペルム紀の祖先については不明な点が多いが、三畳紀の最初期に『主竜形類』(注149)が繁栄したことは明確である。カルー砂漠においては、ペルム紀から三畳紀への移行期と思われるわずか数メートルの地層から、『プロテロスクス(別名カスマトサウルス)』という大型の爬虫類の化石が頻繁に発見されている。これは陸上動物で、その特徴として鋭く尖った歯が列挙される。間違いなく捕食者であり、ワニ類よりは体が地面から少し持ち上がっていたが、脚はワニのように胴体から横に生じていた。
しかし、三畳紀が進展するにつれて、「主竜形類」は体がさらに起きた姿勢へと急速に変化し、細身で動きの速い捕食者が「プロテロスクス」のような初期の主竜形類を置き換えていった。その進化は、速さが要求されたことが一因であるだろうが、同等に重要なことは移動しながら呼吸が可能となる点であったと考えられる。プロテロスクスはトカゲのように、まだ歩行時に体を左右に曲げる動きをしていた可能性がある。このような動きは『キャリアの制約』(<物語り その12>参照)により肺や胸部が圧迫され、歩行中は呼吸が困難になる。トカゲやサンショウウオが歩行しながら呼吸が困難であるのはこのためで、プロテロスクスもまた、ある程度は『キャリアの制約』を受けていた可能性がある。
(注149)『主竜類』にリンコサウルス類、トリロフォサウルス類、原始竜類などの絶滅爬虫類を加えた分類上の単位。
この問題を解消するためには、脚を胴体の下側に配置することが一つの解決策であった。しかし、それだけでは問題は全て解決されなかった。
姿勢と呼吸の問題から完全に解放されるには、運動系だけでなく呼吸器系も大幅に改造する必要があった。
- 恐竜や鳥類の系統は、呼吸の問題を解決するために革新的で効果的な方法を見つけた。『二足歩行』である(図167参照)。四肢動物特有の姿勢を捨てることで、運動と肺機能の制約から解放された。
- 哺乳類の祖先もいくつか新機軸を取り入れている。たとえば、『二次口蓋』(注150)が形成されたことや、四肢動物にもかかわらず、体が完全に起きた姿勢になったことなどである。しかしそれらの改革でもまだ完全ではなく、哺乳類は新たな呼吸器系を進化させた。『横隔膜』という筋肉を使用して強力に空気を吸引・排出できるようにしたのである。
(注150)「口蓋」とは脊椎動物が口腔と鼻腔を分離する口腔上壁のことを指し、「二次口蓋」とは爬虫類までの単層の口蓋から、軟口蓋の先にある口蓋垂(ノドチンコ)が飲み込む瞬間に跳ね上がり気道を閉じることで、二層構造(下層が食物摂取用の口腔、上層が酸素摂取用の鼻腔)を実現したものである。これにより摂食と呼吸が同時に可能になる。
三畳紀の低酸素期における生物の生活状況や直面した困難を知る手がかりとなるのは恐竜の骨だけではない。
三畳紀爆発の特徴の一つとして、海に戻った爬虫類の多様化が挙げられる。多くの系統が海に戻ったのは、三畳紀特有の「高温で低酸素」の環境に対応できなかったためだった可能性がある。
動物の体内での代謝反応には酸素が必要である。代謝は生命活動そのものと言える。しかし、化学反応と同じく、代謝反応の進行はいくつかの要素によって影響を受ける。その中でも「温度」は特に重要である。
『代謝率』とは、生物がどれだけの速さでエネルギーを消費するかを示し、外温動物よりも内温動物のほうがはるかに高い。しかし、同一の生物個体でも、代謝率は温度によって大きく変化し、その影響を直接的に受ける。
近年の研究によれば、動物の一個体が消費する全エネルギーの約1/3から1/2が、生命を維持するためだけに使われている。具体的には、タンパク質の代謝循環(合成と分解の繰り返し)、イオン輸送、血液循環、呼吸などの活動である。さらに、移動や生殖、摂食などの活動もエネルギーを消費し、エネルギー消費率は温度上昇とともに増大する。しかしながら、生物が行う化学反応は酸素がなければ成り立たない。そのため、代謝率が上昇すれば酸素の必要量も増大する。
- この研究から、温度が10度上昇するごとに代謝率が2~3倍になるという重要な結果が得られた。三畳紀では、現代よりも酸素濃度が低いながら平均気温が高かったため、生命活動は深刻な影響を受けた可能性がある。
大気中の酸素濃度と気温との間には直接の関連性は存在しない。しかし、大気中の二酸化炭素濃度と気温との間には直接的な関連性が認められる。これは、いわゆる「温室効果」である。そして地球化学の分野では、大気中の酸素濃度と二酸化炭素濃度が大体反比例の関係にあることが既に注目されてきた。つまり、酸素の量が多い場合、二酸化炭素の量は少なく、その逆もまた真である。過去の低酸素環境では、二酸化炭素の濃度が高く、その結果、温度が高い環境になることが多かった。
気温が高く酸素が少ない状態は、動物にとって不利である。これに対処するためのさまざまな手段が考えられてきた。
- その一つには、体温を最低限の程度にまで下げるという方法がある。これには生理機能の調整、行動の変更、もしくは、体の形態、生理機能、行動の三方面を同時に変化させるという方法も存在する。それが低温の海へ帰ることである。どれだけ気温が高くても、陸上よりは海のほうが低温で生理機能を営みやすいはずである。
- おそらくはこの理由のために、地球の気温が高く(平均気温はたぶん今より約17℃高かった)、大気中の酸素濃度が今日の半分しかなかったこの時期、海の暮らしへと進化しなおした四肢動物はかなりの種類にのぼった。
- あとにも先にもこれほど多くの系統が陸を捨てて海に向かった時代はない。中生代の陸上動物のかなりの割合が足をヒレ足やヒレに変えて海へと戻っていったのである。

現代でもクジラやアザラシ、ペンギンのように、元々は陸地に生息していた動物が海へと適応し、多様な種を形成している。ただし、哺乳類全体の中でクジラやアザラシの種が占める割合は2パーセントに過ぎず、ペンギンは鳥類全体の1パーセントを占めるに過ぎない。しかし、三畳紀の海には、現在よりも多くの、陸の体制から海の体制に変化した動物の種が存在していた。この時期には巨大な「魚竜類」(図168参照)や、「板歯類」(図169参照)など、海に回帰した四肢動物も見られた。板歯類は大型のアザラシのような姿をしていたが、アザラシと異なり、歯が尖っていなかった。これは主に貝類を食べるために進化した結果である。
ジュラ紀にはまだ魚竜類が残存していたが、首長竜類も多数見られた。白亜紀になると魚竜類は消え、大型のモササウルス類がそれに取って代わった。

これら全てに共通するのは『海への回帰』というテーマである。その時代の大きな環境要因としては、酸素濃度の低さと世界的な高温が挙げられる。
ワシントン大学で爬虫類を専門に研究するレイ・ヒューイ博士も、三畳紀前期からジュラ紀を通して気温が高かったことが、多くの爬虫類が海に回帰する動機になったと指摘している。
さらに、ピーター・ウォード博士は2006年に、中生代の酸素濃度と海生爬虫類の数との間に興味深い反比例の関係があることを示した。その結果、
- 酸素濃度が低い時には、海生爬虫類の科が全体に占める割合が大きかった。
- しかし、酸素濃度が高くなると、完全に海生の四肢動物の科が占める割合が著しく減少した。
- これは、海洋動物の絶対数が減ったというより、陸上の恐竜の数が急増した結果である可能性がある。
三畳紀の黒色
三畳紀の黒色地層は、原因が明確でない『三つの大量絶滅イベント(ペルム紀、三畳紀、暁新世)』(<科学物語り その10>図113参照)が発生した時に、それぞれの地層が低酸素状態で堆積したことを示している。低酸素状態では地層は一般に黒色になる。この「黒色頁岩」と称される岩石は黄鉄鉱や硫黄化合物を含むが、これらの物質は通常の酸化環境の海洋では分解し、保存されにくい。したがって、黒色頁岩は海底が貧酸素~無酸素状態になった時に堆積した物質であることが示唆される。
二つ目の手がかりは、それぞれのイベントが起きた時期の地層が薄層状であり、場合によっては薄板状になっていることで、繊細な堆積構造が見られることが多いという事実である。カンブリア紀以降、穴を掘る動物が多数出現し、海底に堆積した地層のほとんど に生物による擾乱の痕跡が残っている。多数の無脊椎動物が海底の堆積物を体内に取り込んで有機物を濾し取る結果である。地層が薄層状に堆積している場合、その環境には動物がほとんどいなかったか、あるいは数が極端に少なかったと推測される。
以上のように、
- 酸素濃度モデル、岩石の色、そして堆積物の層構造という三つの観点から考えると、「ペルム紀、三畳紀、暁新世の絶滅が低酸素世界で起こった」という結論に至ることは明らかである。
1990年代後半から21世紀の初めにかけてはほかにも証拠が発見され、酸素とは対照的に「二酸化炭素」という別の大気成分が高い濃度だったことが確認された。低酸素の場合と同様、「二酸化炭素濃度が高かった」という証拠もバーナーのモデルから得られ、さらには岩石の記録から、もっと正確に言えば化石記録からも裏付けられている。
残念ながら、どの年代においても、実際の二酸化炭素の量を正確に測定する手段は存在しない。二酸化炭素は岩石に色をつけることもなく、層状構造に影響を及ぼすこともない。しかし、巧妙な方法で葉の化石を分析した結果、二酸化炭素の相対的な量を推定する突破口が開かれた。この方法を用いることで、二酸化炭素が増加していたのか、減少していたのか、または変動していなかったのかを百万年単位で把握することが可能となり、基準値から何回上下動したかも推測できる。
具体的にはどのような手法なのだろうか。この二酸化炭素濃度の測定法は巧妙かつ単純なものであって、現代の植物の葉を研究している植物学者が、閉鎖系で植物を育てる実験を実施し、現在の大気中濃度(最初にこの実験が行われた時点では約360ppm)と比べて二酸化炭素を増やしたり減らしたりできるようにしたのである。植物は二酸化炭素濃度に非常に敏感であることがわかっている。大気に含まれる二酸化炭素がどんなに少量であろうと、それが植物の炭素源、生命を構成する重要な要素であるからだ。
- 二酸化炭素は主に葉を通じて取り込まれ、その過程は微細な孔である『気孔』を介して行われる。高濃度の二酸化炭素環境下で育てられた植物は、生成する気孔の数が減少する。それは、二酸化炭素が豊富に存在すると、少ない数の気孔でも十分であるからだ。そこで植物学者たちは、化石記録を詳しく調査することに取り組み始めた。
- 葉の化石では気孔の数が簡単に観察できる。結果はバーナーのモデルを裏づけるものだった。
- ペルム紀末と三畳紀前期の間では、葉の化石にはわずかな気孔しか観察されなかった。結果として、「三つの絶滅において、二酸化炭素の濃度が驚くほど高かった」ことが確認された。しかも、その濃度上昇は数百万年間ではなく、わずか数千年間で起きていた。
この二つの結果から見えてくるのは、大量絶滅の新たな姿である。
- 「三つの絶滅が起きたときには、短期間に急激に増えた二酸化炭素によって(他の証拠によれば恐らくメタンによっても)世界が急速に温暖化した。
- 暑さと酸素欠乏が重なった。
- この高温低酸素の時期は、大規模な大量絶滅が発生した時期と一致している。
暑さは動物にとって大敵である。動物が耐えうる最高気温は、水が沸騰する温度の半分にも達しない。40℃ではほとんどの動物が死ぬし、45℃になれば生存は困難となる。さらに、温度上昇が生命に危険を及ぼす可能性は、車内に子供が放置されている悲劇的な事件からも明らかである。
さらに、熱エネルギーの量と利用可能な酸素の量は生命の機能にとって重要で、動物が必要とする酸素量は気温上昇に伴って増加する。したがって、高温と酸素欠乏が重なれば、生存環境は極めて厳しくなる。
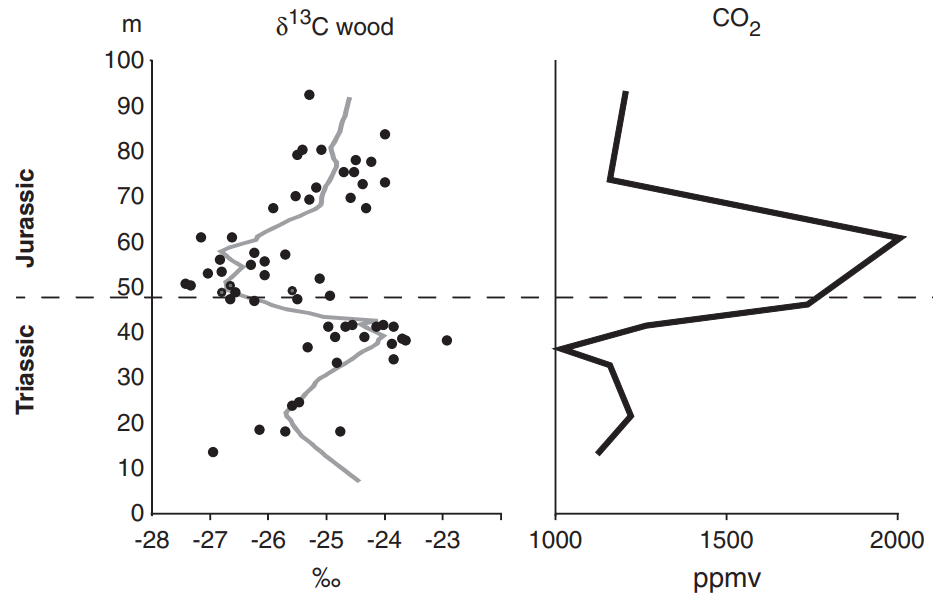
三つの絶滅の中でも、「T-J境界(三畳紀とジュラ紀の境目)」の二酸化炭素増加を示すデータ(図170、注151)は特に注目に値する。シカゴ大学の古植物学者ジェニファー・マケルウェインは、20世紀末頃、氷に閉ざされたグリーンランドの危険な凍土層からサンプルを収集した。
(注151)Margret Steinthorsdottir, Andrew J. Jeram and Jennifer C. McElwain, “Extremely elevated CO2 concentrations at the Triassic/Jurassic boundary,” Palaeogeography, Palaeoclimatology, Palaeoecology 308 (2011)418-432.
その結果、
すでに低酸素だった世界で二酸化炭素が急上昇したことが、三畳紀の終わりを決定付けたものと疑問の余地なく確認された。徐々に、三畳紀末の事象はペルム紀末のそれと似ていると認識されるようになった。
- 『K-T境界(白亜紀と第三紀の境界)』の絶滅イベントとは明らかに異なる特徴を三畳紀末の絶滅イベントは示している。K-T境界においては、突如として絶滅が起き、あらゆる動植物に影響が及んだ。それに対する予兆は、生態系の側面でも進化の側面でも何一つ見られなかった。
- それに対し、三畳紀末では竜盤類を除く恐竜群がT-J境界前後の時期に小型化の傾向が見られた。まるで、困難な時期が訪れることを予知し、小型化することで適応を試みていたかのような様相であった。
T-J境界絶滅では、最も単純な構造の肺を持つ動物群(両生類や初期の爬虫類)が大打撃を受けた。「植竜類」(注152)を含む、三畳紀前期に繁栄した多くのグループが全滅した。両生類や主竜形類の肺の構造は単純で、肺の膨張はおそらく肋骨の筋肉組織に依存していた。この時期の哺乳類や高度に進化した獣弓類は、「横隔膜」を用いて肺を膨張させていたと推定され、絶滅の被害は比較的少なかった。一方、ワニ類はおそらく腹部のポンプを使って肺を膨張させていたため、大きな被害を受けた。「竜盤類恐竜」が生き延びた背景には、食物の獲得の容易さ、高温への耐性、捕食者の回避、繁殖の成功などの要因が関与していたと考えられる。
(注152)植竜類は地球上にかつて生息していた主竜類の中の一つで、体長4mに及ぶワニに似たフィトサウルスで代表される。ワニと近縁で体型も似ているが直接の祖先ではない。恐竜、翼竜、鳥類などとは遠い類縁関係にある。現在のワニのように水中生活を送るものが多かったが、陸上生活に適応した種も存在していた。
注目すべき点は、「竜盤類」だけが高度な「隔壁式の肺」を持っていたという事実である。
- この肺は、内側の表面積を増加させるために多くの小さな隔壁組織を有し、他の系統の動物の肺よりも効率が良かった。T-J境界絶滅の前後では酸素濃度が極端に低かったため、このような呼吸器系を持っていたことが生存に極めて有利だったと考えられる。
- もし上述のシナリオが正しければ、竜盤類が三畳紀末に覇権を握り、ジュラ紀に入ってからも支配的な地位を保持できたのは、「優れた肺」を持っていたことにより、活動レベルが他の生物よりも高かったからと推定できる。
- 三畳紀の中期から後期にかけては、多様な体制の爬虫類が登場したが、その中で竜盤類だけが多様化したことが現在では明らかとなっている。
他のグループは多様性が変わらないか、あるいは低下したかのどちらかで、後者が多かった。三畳紀後期には酸素濃度が過去5億年の中で最低レベルに落ちたことが明らかになっている(図114参照)。竜盤類が持つ特性の何らかが、低酸素の環境でも生存する力を向上させたと考えられる。地層の調査結果からは、酸素濃度が徐々に低下し、最終的に三畳紀の大量絶滅に至ったように見える。また、その絶滅が実際には一度ではなく、300万~700万年の間隔をあけて二度起きたと推測される(注153)。
(注153)三畳紀の終わりに、再びやや小規模な大量絶滅が起きた。海洋ではアンモナイトの多くの種が姿を消し、魚竜などの海洋棲爬虫類も大打撃を受けた。陸上ではキノドン類、ディキノドン類の大部分の種といった大量の単弓類(哺乳類型爬虫類)が絶滅した。三畳紀の終末を生き延びた恐竜など陸生脊椎動物は、繁殖様式(卵など)や生活様式から乾燥にとくに強いタイプのものと推定される。また、爬虫類も単弓類同様に大型動物を中心に多くの種が絶滅した。まだ比較的小型だった恐竜は、三畳紀末期には竜脚類のような大型種も出現し、その後急速に発展していった。
この時期の地層で、陸生脊椎動物の化石が豊富に見つかるような場所はほとんど存在しなかった。そのため、陸生脊椎動物がどのように絶滅していったかは、海洋生物と比べて詳細が明らかになっていないのが現状である。大量絶滅の犠牲者として知られる動物たち(植竜類、アエトサウルス類、原始的な主竜形類、獣弓類の一種トリティロドン類などの大型動物)が、どれだけの期間で姿を消したのかも確認できていない。しかし、
華麗なジュラ紀のアンモナイトが海に大挙して現われ、ジュラ紀初期の岩石に彼らの復活の痕跡を豊富に残す時期には、『恐竜』がすでに世界を掌握していたことは明らかである。
では、恐竜の肺はどのようなものだったのだろうか。確実にいえるのはただ一つ。
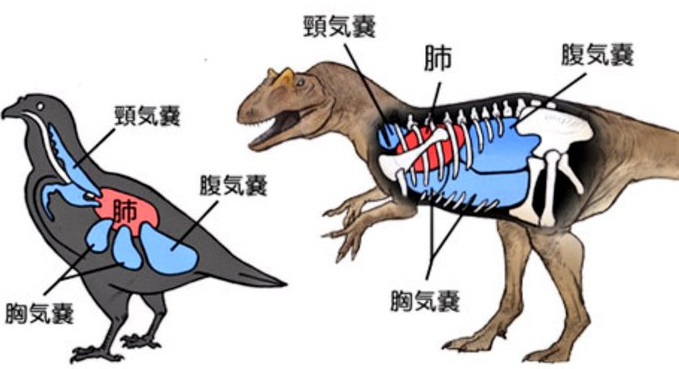
- 「動物の誕生以来最悪となった酸素危機にも、見事に対処できる肺や呼吸器系を備えていた」ということである。
陸生脊椎動物の中で、竜盤類恐竜の絶滅率が最も低かったのは、
- 競争に勝つための高度な呼吸器系、つまり『最初の気嚢システム』を持っていたからであると、新たな視点(注154)で考えられている。
- 竜盤類がT-J境界の大量絶滅期を経てむしろ数を増やしていたという事実は、何より特筆すべきことである。
(注154)恐竜が初期の哺乳類を生存競争で圧倒したのは、肺の機能が優れていたことが原因であるとする研究が発表されている(注155)。
(注155)C. G. FARMER AND KENT SANDERS,“Unidirectional Airflow in the Lungs of Alligators”, Science 327 (2010) 338-340.
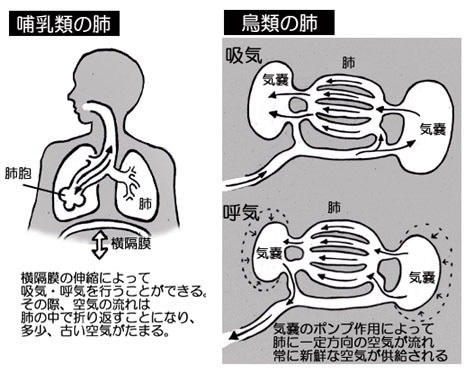
恐竜と共通の祖先を持つ現生のアリゲーターを研究したところ、鳥類と同様の高効率の呼吸法を持っていることが明らかになった。哺乳類の場合、息を吸うたびに酸素を豊富に含んだ空気が「肺胞」と呼ばれる肺の中の“袋”に運ばれる。肺胞で空気が循環することにより、酸素が血流に運ばれ、血液中の不要な二酸化炭素が排出される。しかし鳥類には肺胞が存在せず、空気は「気嚢」に一方通行で流れ込むように進化している。その結果、鳥類の肺は常に新鮮な空気で満たされる(図172参照)。この機能により、鳥類は他の動物が生存できない高所でも呼吸することが可能である。今回の研究では、アリゲーターの呼吸法を明らかにするために、死亡したアメリカアリゲーターの肺に液体を注入し、空気の流れる方向を調査した。その結果、鳥類と同様に、吸った空気が体内に流れ込む際には特定の気管支の層を迂回し、呼吸時に初めてこれらの気管支を通過することが確認された。
このような呼吸法は、2億5100万年前から1億9900万年前の三畳紀に生息していた、鳥類、恐竜、アリゲーターの共通の祖先にも見られた可能性が高い。鳥類、恐竜、アリゲーターは同じ主竜類に分類されており、三畳紀初期には大気中の酸素が現在よりも希薄であった。
図表
図162 スミシアン末期に発生した最も高温な環境
高橋 聡Res.Org.Geochem. 29 (2013) 1−16,“ペルム紀-三畳紀境界の遠洋域深海堆積岩の
古生物学的地球化学的研究”
図163 腕足動物の模式図
フリー百科事典『ウィキペディア(Wikipedia)』「腕足動物」(https://ja.wikipedia.org/wiki/%E8%85%95%E8%B6%B3%E5%8B%95%E7%89%A9)
図164 アンモナイト亜網の各種化石
アンモライト研究所/アンモナイトの進化について
(https://www.ammolite.co.jp/evolution/index.html)
図165 三畳紀の恐竜
iStock by Getty Images(https://www.istockphoto.com/jp/%E5%86%99%E7%9C%9F/%E4%B8%89%E7%95%B3%E7%B4%80)
図166 プロテロスクスの想像図
『ウィキワンド(Wikiwand)』「プロテロスクス」(https://www.wikiwand.com/ja/%E3%83%97%E3%83%AD%E3%83%86%E3%83%AD%E3%82%B9%E3%82%AF%E3%82%B9)
図167 テコドントサウルスの想像図
『ウィキワンド(Wikiwand)』「テコドントサウルス」(https://ja.wikipedia.org/wiki/%E3%83%86%E3%82%B3%E3%83%89%E3%83%B3%E3%83%88%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9)
図168 イクチオサウルスの想像図
『ウィキワンド(Wikiwand)』「イクチオサウルス」(https://ja.wikipedia.org/wiki/%E3%82%A4%E3%82%AF%E3%83%81%E3%82%AA%E3%82%B5%E3%82%A6%E3%83%AB%E3%82%B9)
図169 プラコドゥスの想像図
古世界の住人
(http://paleontology.sakura.ne.jp/purakodosu.html)
図170 T-J境界の二酸化炭素増
Margret Steinthorsdottir, Andrew J. Jeram and Jennifer C. McElwain, “Extremely elevated
CO2 concentrations at the Triassic/Jurassic boundary,” Palaeogeography,
Palaeoclimatology, Palaeoecology 308 (2011)418-432.(https://www.academia.edu/23075570/Extremely_elevated_CO2_concentrations_at_the_Triassic_Jurassic_boundary?from=cover_page)
図171 恐竜と鳥の気嚢システム
古世界の住人
(https://ameblo.jp/oldworld/entry-10153482359.html)
図172 哺乳類・鳥類での呼吸システム内の空気の流れ
古世界の住人
(http://stat.ameba.jp/user_images/3a/49/10102504261.jpg)

{kind=link}