
第四部 生命の進化
第7章 動物の発展【4億7500万 ~ 3億年前】
1.動物の陸上進出【4億7500万 ~ 3億年前】
海洋動物の分類群(種、属、科など)がカンブリア爆発以降増加していることは、簡素な検証を行うことにより確認された。具体的には、『生痕化石』の数の推移を調査したのである。
カンブリア爆発についての章で触れたように、生痕化石は動物が行動した結果として生じる。生痕化石の種類が異なれば、それを残した動物の形態も異なると考えられる。つまり、生痕化石の多様性は体化石の多様性を反映するものである。古生物学者が古代の無脊椎動物を研究する際には、生痕化石にさまざまなパターンがあることに注意してきた。現在では、生痕化石が地球上での生命の多様化の経緯を明確に示す手段となっているというのがこの分野の共通の理解である。
デボン紀が終わる頃には、海は浅瀬から深海に至るまで、生物でほぼ埋まるほどになっていた。しかし、この海洋生物の多様化さえも、もうすぐ起ころうとするさらに大規模な出来事の前では影が薄くなる。それは、地球上で最も大量の動植物の種を生み出す「陸上における生命の多様化」である。
進化論に批判的な視点からは、『失われた環(ミッシングリンク)』(注99)が存在しないのではないかという指摘が常になされてきた。しかし、21世紀に入っての新たな発見がこれを覆すこととなった。
(注99)生物の進化過程を連続した鎖と見なした場合、その鎖の中に存在すると考えられるが見つかっていない部分(間隙)を指す。祖先群と子孫群の間に存在すると予想される進化の中間期に当たる生物・化石が見つかっていない状況を指す。
厳寒のカナダ北極圏のデボン紀の地層から、特筆すべき化石が発見された。それは『ティクターリク』と名付けられ、その特徴が魚類から両生類への過渡期を示すものであったため、発見者たちはこれを『足を持つ魚』と呼んだ。水生脊椎動物から陸生脊椎動物への移行に関する問題は大きなギャップを残していたが、この発見はその問題を解決するだけでなく、進化論そのものをより堅固なものとする一助となったのである。

この大型化石はシカゴ大学のニール・シュービン率いる国際的な研究チームによって、カナダ北極圏地方で発掘された。苦心の末にようやく石棺のような堆積岩の覆いが取り除かれたとき、最初は一見するとただの魚かと思われた。鱗(うろこ)も鰓(えら)もあり、頭部は平らで、鰭(ひれ)には細い鰭条(きじょう)骨(注100)があり、普通の魚のひれと区別がつかない。しかし、この新種の魚は、内部に頑丈な骨格を持っていた。この骨格のおかげで、体長90センチにも及ぶ大きな動物でも、四肢に似たひれを支えにして浅瀬から身体を起こすことが可能であった。
(注100)魚類の鰭(ひれ)を構成する膜状部を支える細長い骨で、基部は担鰭骨に支えられ、鰭の外縁に向かってのびる。棘(きょく:触ると固く先端が尖っている)と軟条(魚のひれにある軟らかいすじ)の二種がある。
この奇妙なひれと、両生類的な(ワニに似た)頭部を併せ持つ「ティクターリク」は、魚の体制から四肢動物の体制へと段階を追って進化していったことを示す絶好の例といえる。
次々と海洋動物と植物が陸上へと進出していった。脊椎動物の初の陸上進出は最も劇的な出来事であり、人間にとっても直接的な関係がある。しかし、実際には、私たち脊椎動物が陸上に出たのは最後のことであった。それ以前にも、多くの動物が水から陸上への移行を達成していた。まずは、最初に水から出てきた生命体、植物から話を始めよう。
植物の上陸
生命の誕生を除けば、生命の歴史における最も重要な出来事は、生物が「酸素発生型光合成」を発明したことであると言えるだろう。
太陽系最大のエネルギー源である太陽のエネルギーを利用できるようになったことで、生命は暗く湿った場所から脱出し、浅瀬を満たし、淡水環境にも進出することができた。その過程で、意図しない副産物が生じた。地球の大気は大きく変化し、酸素濃度が上昇したのである。その結果、予期しない結果として、植物を食べる動物が登場し、植物生命にとって大きな脅威となった。こうして、
- 水生植物が生命に及ぼした影響は非常に大きかったが、植物自体にも大きな変化が訪れることになった。それは、水の制約を乗り越えて乾燥した大地で生きるための手段を身につけることであった。
- 地球の歴史の中でわずかな時間、全体の1パーセントにも満たない期間に、植物の陸上進出は全てのルールを変え、生命の歴史を新たな方向へと導いたのである。
現在把握されている事実を総合すると、以下のようにまとめることができる。
- 海に生命が誕生してから約1億年以内に、淡水域に生息していた可能性が高いある種の『緑藻類』(注101)が水中の生活から離れ、陸地へと進出した。
- 初期の段階では葉を持たず、小枝のような形態であり、現代のコケ類に似た姿だった。しかしこれらの生物は急速な進化を経験し、巨大な植物へと進化した。その主要な要因は、進化が生み出した新たな構造、すなわち『葉』の登場である。

(注101)緑藻類とは、体が緑色で、光合成の主要色素としてクロロフィルaとb(クロロフィルaが植物や藻類の光合成の主成分であるのに対し、クロロフィルbはエネルギーを集めてクロロフィルaに渡す副成分である)を保有し、光合成によりデンプンを生成し貯蔵する緑色植物のうち、維管束を持つ種子植物、シダ植物、胚を持つコケ植物、多細胞の生殖器官を持つ車軸藻類を除いた残りを指す。全世界で約1万種近くが知られ、そのうち90%近くが淡水産で、残りの10%弱が海産である。
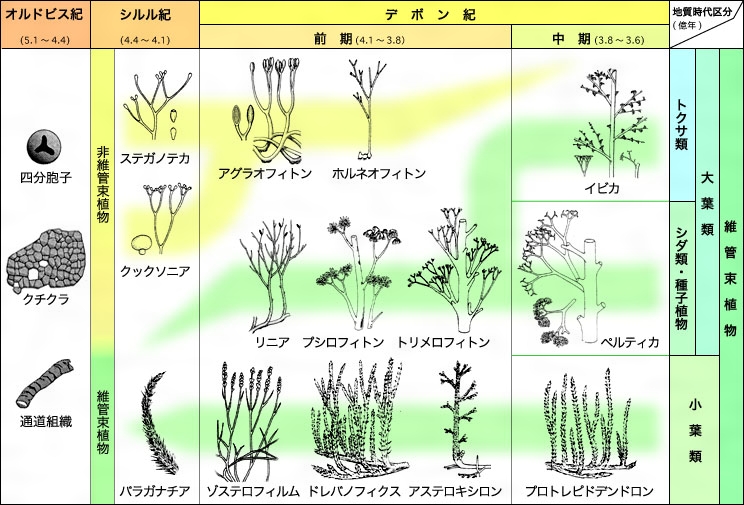
- 約4億7500万年前から、水生だった緑藻類は様々な構造を進化させ、水だけでなく、空気と土の組み合わせからも養分を得られるようになった。この過程で最も重要だったのは、このような環境での生殖が可能となったことである。
- 約4億2500万年前には、間違いなく『維管束植物』(根と茎を持つ)の化石が初めて出現した。その間の詳細は化石記録には残されていないが、必要な変化を経ながら進化は一歩一歩進行していたのである。
- そこから本物の「葉」を持つ植物が誕生するまでには、さらに4000万年を要した。
- しかし、一度最初の葉が現れると、革新的な変化が迅速に起こった。約3億7000万年前から3億6000万年前の間には、木々が既に高さ8メートル近くにまで達していたのである。
- 上陸した多細胞植物は、約1億年かけて小型の海洋植物の形態から樹木へと姿を変え、デボン紀の終わりには森林が地球を覆うまでになった。
- 陸地を長らく支配していた微生物に比べ、陸生の多細胞植物が地球に及ぼした影響は遥かに大きかった。陸生の多細胞植物によって、地形と土壌の性質が大きく変わり、大気の透明度も変化したのである。
それまでの地球では、砂丘が絶えず移動し、砂塵が舞っていたが、陸上に広がる植物の数が増えると、その状況は劇的に変わった。微生物に比べ、植物の根は砂や塵をより強固に保持できる。これは単細胞の微生物や、細菌のマットでさえも到底及ばない能力である。原始的な植物が枯れ、その場で分解されることにより、土壌が形成され、厚みを増し、かつての岩石だらけの荒涼とした風景は次第に和らいでいった。この変化は、宇宙から見ても明らかであった。大気が澄んだことで、大陸や海、そして大きな湖や川の輪郭がはっきりと見えるようになったのである。
- デボン紀後期に入ると、森林が陸地をほぼ覆い尽くし、川の流れそのものを変えるまでになった。
- この期間中、植物は大気中の酸素濃度を引き上げ、現代の21パーセントを大幅に上回る30~35パーセントまで増加させた。このような高濃度の酸素環境ならば、肺を持たない四肢を持つ魚でも海から這い出て、効率的に空気を呼吸する肺に進化するまで生き延びることが可能であったと考えられる。陸上植物がもたらしたこれらの変化は、すべて革新的な構造、「葉」の誕生に起因するのである。
陸と海の異なる環境-動物が海を離れた理由
陸地と海は異なる環境である。動物が海から陸地に一斉に移動したわけではなく、少数ずつ、何度かに分けて上陸を試みたが、その過程で多くが死亡した。では、なぜ最初に動物が海を離れたのか。最近の説では、従来の説明、つまり、
- 陸には未開の資源が存在し、競争が少なく、捕食者も少なく、動物自身も上陸できる段階にまで進化したから
というものとは異なり、
- 最初の動物の上陸を可能にしたのは『大気中の酸素濃度が上昇したから』だ
という見方がされているのである。
まず、動物と植物が共に陸上生活を営むためには、何が必要かを考えてみると、
- 初期段階での植物の存在が重要である。これは、植物が食糧源としての役割を果たすため、その存在がなければ動物は陸地に生息する足場を築くことが困難である
からである。
約6億年前、植物は様々な系統の多細胞生物へと多様化を遂げていた。現代でも海岸にて観察できる『緑藻類』や『褐藻類』、『紅藻類』(注102)などもその一部である。
とはいえ、これらは、海水の中で進化した植物である。生存に必要な要素、つまり二酸化炭素と養分は、周囲の水から容易に摂取できた。また、生殖は液体の環境を介して行われていた。
- 陸上への適応は、二酸化炭素や養分の取得、体の支持、生殖の各面で、進化による体の構造の大きな変化を必要とする。完全に水生の生物にとって、これらのどれ一つにおいても既存の体制の大幅な改革が求められる。
最初の植物の上陸については未だに明確な知見が得られていない部分が多い。特に原生代において、スノーボールアースが発生する前にどれだけの植物がどの程度多様に存在していたのかは、未解明の謎である。
(注102)『緑藻類』・『紅藻類』・『褐藻類』という三つの藻類グループの植物としての具体的な特徴の違いとしては、まず、光合成を行う細胞小器官である「葉緑体」の内に含まれる色素の種類がある。「緑藻類」の場合は、通常の陸生植物と同様に緑色の色素である葉緑素クロロフィル(葉緑素)が主体となる。一方、「紅藻類」の場合にはクロロフィル(葉緑素)の他に、フィコエリトリン(紅藻素)と呼ばれる赤色の色素や、フィコシアニン(藍藻素)と呼ばれる青色の色素も多く含む。そして、「褐藻類」の場合は、クロロフィル(葉緑素)とともに、フコキサンチン(褐藻素)と呼ばれる赤褐色の色素が多く含まれている。また、「緑藻類」には海藻だけではなく淡水産の藻類も数多く含まれるのに対して、「紅藻類」と「褐藻類」に属する藻類はそのほとんどが海産の藻類であり、そうした海藻の中でも、「緑藻類」の場合は比較的水深の浅い海域に生息する藻類の種類が多いのに対して、「紅藻類」には深海などの水深の深い海域に生息する藻類が数多く分類され、「褐藻類」には全長が数メートルから数十メートルにも及ぶような大型の藻体を形成する藻類の種類が数多く分類されるといった点が「緑藻類」・「紅藻類」・「褐藻類」の三者を区別する主要な特徴の違いとして挙げられる。
光合成生物であり、「植物」と呼べる体制と多様性を持つ存在は、約4億7200万年前よりも遥か以前から多く存在していた。多種多様な『真菌類』(注103)や、光合成をする緑色の微生物、さらには多細胞の植物まで、定説より早く上陸していたのではないかと考える古生物学者は多い。また、『地衣類』(注104)や『真菌類』、あるいは湿地や沼地を覆う緑色の『微生物マット』を植物の範囲に含めるなら、すでに10億年前には植物と総称できる生物の群集が形成され、驚くほど繁栄していた可能性がある。
(注103)真菌類はほとんどが固着性であり、キノコ、カビ、単細胞性の酵母、鞭毛を持った遊走子などの多様な形態を示す真核生物で、「菌界」に分類される。大部分の真菌類は、外部に分解酵素を分泌して有機物を消化し、細胞表面から摂取する従属栄養生物である。
(注104)地衣類は、藻類(シアノバクテリアあるいは緑藻)を共生させることで自活できる菌類である。一見ではコケ類などに似て見えるが、形態的にも異なり、構造は全く別のものである。

しかしながら、「本物の植物」と認識され、通常、生命史における最古の植物として認められているものは、最終的に緑藻類の一種である『車軸藻類』(注105)から派生したとされている。
(注105)車軸藻類 (しゃじくもるい)は、水草のような形態を持つ大型の藻類で、輪生(生物の体において、ある器官が一定の箇所から輪を描くように並んで生じること。植物の葉や花に関して使われることが多い)する小枝や節と節間の繰り返しからなる特徴的な体と、極めて複雑な生殖器官を有している。多くは淡水止水域に生育するが、特に湖沼では比較的深い水深まで分布している。
そこへ到達するには、多数の障害を乗り越えねばならなかった。
- 特に最初に解決すべき課題は「乾燥の防止」であった。
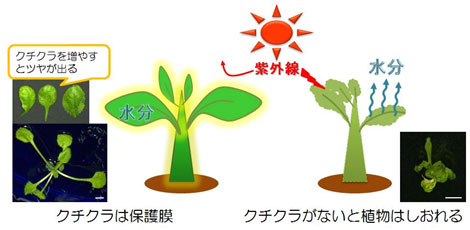
緑藻類は水中生活者であるため、海浜に打ち上げられると急速に脱水して死滅する。体を守るコーティングが存在しないため、すぐに脱水状態となる。しかしながら、車軸藻類の「接合子」(注106)は、防水性を持つ『クチクラ』(注107)で覆われている。このクチクラが、陸上進出時に植物全体を覆う役割を果たした可能性がある。だが、クチクラの発達は新たな課題を引き起こした。それは、二酸化炭素の取得が困難になったことである。海中では二酸化炭素は水に溶け、細胞壁を通じて吸収するだけでよかったが、陸上に適応した初期の植物は、『気孔』という微細な穴を多数形成し、そこから二酸化炭素を取り込む方法を開発した。
(注106)「接合子」または「接合体」は、配偶子が接合する結果形成される細胞である。例えば受精によって形成される受精卵もその一例である。接合は有性生殖の一形態であり、接合子はその結果生じる細胞で、新たな個体へ発展する。通常、2つの半数体の細胞が合体し、接合子または接合体と呼ばれる二倍体の細胞を形成する。
(注107)クチクラは植物の表面に光沢を与える脂質ポリマーで、最表面に形成され、風雨、乾燥、紫外線、病原菌などの外部環境から植物を保護している。
- 初期の陸上植物が体を地面に固定する際には、『真菌類』(注103)との共生を利用していたと考えられる。これは固定器官が確認できないためである。また、この共生関係は土壌から水分を得る上でも有用であった。
- 陸上進出に伴い、体を支える方法についての問題も生じた。
植物は日光を吸収する表面積を可能な限り広げる必要があり、最初の陸上植物は恐らく地面に密着する形でこれを達成した。今日でも、コケ類はこの方式を用いて、土地をカーペットのように覆う。オルドビス紀の陸地に存在したとされる大きな「木」であっても、その高さはせいぜい5〜6ミリであった。しかし、この方法には限界がある。垂直方向に成長すれば、より多くの日光を獲得することが可能である。地面に広がるタイプの植物間で競争が生じる生態系では、これは特に重要である。したがって、
- 初期の植物は様々な硬質物質を組み込んで初の茎を生み出し、最終的には木の幹へと発展させた。
- それと同時に、新たに形成された根から葉への輸送システムも進化した。
- 最後に、乾燥に耐える生殖体を作り出し、陸地環境で確実に生殖できるようにした。
これらの新しい進化により、植物は陸地に定着した。その結果、大量の有機炭素が初めて陸上に形成され、動物がすぐに続いた。新しい資源は新しい進化を引き立てる。大半の見解では、初期の陸上植物は淡水性の緑藻類から進化したとされるが、化石記録でその証拠を確認するのは困難であった。存在する化石は極めて断片的なもののみであり、これらの化石の発掘作業は、文字通りと比喩的にも、探偵のような優れた推理を必要とした。
陸上植物は、陸上の生態系を支配する過程で、地球の気温や海洋の化学的性質、そして大気の組成を変化させ、地球上の生命の性質を変えた。

その歴史を振り返ると、ウィリアム・ランダーという古植物学者は、ウェールズ地方で最古とされた陸上植物の化石を発見した。その化石が発見された岩石は約4億1700万年前のものであった。その後、ウェールズ地方の岩石から、より古い約4億2500万年前の化石が確認された。
この最古の陸上植物は『クックソニア』(注108)と命名され、それ以降の植物は驚くほど長い時間をかけて分布を拡大した。
- 4億2500万年前~3億6000万年前の時代には、植物版のカンブリア爆発が起きた。
- しかし、最新の見解によれば、陸上植物が最初に登場してから少なくとも3000万年のあいだは『葉』は存在しなかった。葉を持つ植物が定着したのはおそらく3億6000万年前以降であると考えられている。
- 葉が誕生するまでに長大な時間が必要だった理由は、依然として未解明である。葉を持つ植物が現れてから、その数と多様性が地球全体に広がるまでにはさらに約1000万年を要した。
(注108)ここで述べている植物は、古生代シルル紀中期からデボン紀前期に生息していた陸上植物の一属である。学名は、オーストラリアの著名な古植物学者イザベル・クリフトン・クックソン(1893〜1973)にちなんで命名された。高さは数センチメートルほどであり、茎の先端にはトランペット型の胞子嚢を持っていたと推測される。地衣類と細菌に続き、初めて海から陸上に進出した生物群の一部であり、乾燥から保護する機能を備えていた。その特徴として、維管束を持たず、シダ植物とは形態が異なる点が挙げられる。また、蝋質のクチクラ層を持ち、植物体内の水分蒸発を防いでいた。しかし、クチクラの防水性は水分を保持する一方、植物内への気体の取り込みを制限するため、ガス交換を効率的に行うための『気孔』が発達したことが化石から確認されている。これらの植物は陸上を開拓した先駆者であり、2011年現在に確認された陸上植物の中では最も古いもの、そして維管束植物の祖先とされる。
陸上植物の進化を理解するためには、エボデボと遺伝子の役割に注目することが重要である。植物は初めに、葉を構築するための遺伝子の道具箱を進化させなければならなかった。その後、この道具箱を上手く利用する必要があり、この過程には時間が掛かったと考えられている。
現時点で最も信頼できるデータによれば、葉を持つようになった植物には葉を形成する遺伝子が存在していたが、その活性化には周囲の環境条件が適切であることが必要だった。
- 動物が高い酸素濃度を必要とするのとは対照的に、植物は大気中の二酸化炭素濃度の低下を待つことが求められていた。
これが、21世紀の古植物学における最新の解釈である。
この理由は現代の事例からも理解できる。生存している植物を実験対象とすると、植物が周囲の二酸化炭素濃度に非常に敏感であることが明らかになる。光合成を行うために二酸化炭素は不可欠であり、その取り込みには周囲の二酸化炭素の吸収が必要である。
葉が存在する場合、外部からの二酸化炭素は葉の外壁を通過し、「気孔」と呼ばれる小さな穴から内部に入る。しかし、この気孔は双方向性を持ち、二酸化炭素が取り込まれると同時に体内の水分も逃がす。陸上動物と同様に、陸上植物にとっても『乾燥』は生存を阻害する大きな障害であり、この問題をどのように解決するかは進化の過程で再三にわたり現れる課題である。
環境中の二酸化炭素濃度が高いときは、気孔の数は非常に少なくなる。逆に、二酸化炭素濃度が低いときは、気孔の数は増加する。二酸化炭素濃度が高い環境は、陸上植物にとって適していると思われるかもしれない。これは生理機能の観点からは正確である。しかし、二酸化炭素は温室効果ガスであり、濃度が高いと地表温度も高くなる。
植物には精緻な信号伝達システムが存在し、成熟した葉から成長途上の葉へ情報を伝えることが可能である。例えば、周囲の環境に適合するためにはどれだけの気孔を形成すればよいか、といった情報を伝達できる。もし私たちが陸上植物が進化を始めた4億年以上前に戻れるとしたら、二酸化炭素濃度が著しく高く、その結果非常に高温の惑星を見ることになるだろう。この高温は、植物の進化と生態系の繁栄に大きな制約を加えていた。気孔は二酸化炭素を取り込みながら植物体内の水分を逃がし、その結果植物を冷却する効果がある。
乾燥は一定の範囲で植物を冷却する効果があるが、その度合いが過度に高いと、体温を大幅に下げる必要がある。しかし、大気中の二酸化炭素濃度が高いときは、植物は少ない気孔数でも二酸化炭素の要求を満たすことができる。しかし、気孔が少なすぎると、体の冷却は十分に行えない。葉のように面積が大きく、表面が平らな部分に気孔が存在する場合、この問題は更に深刻化する。
- 大きな葉がわずかな気孔しか開けていないと、過熱を起こし、葉が死滅する。葉の誕生に時間がかかった理由として、この点が新たな視点であるとされている。
- 遺伝子の道具箱は葉の形成を可能にしていたものの、二酸化炭素濃度が高い大気環境では、植物が積極的に葉を生やすことはなかった。ビアリングらが21世紀初頭に発表した研究は、植物が葉を維持するには二酸化炭素濃度の低下が不可欠だったと示唆している。そのため、植物が葉を持つことは、当時の環境では存続の難しさを増すものであった。
- だからこそ、最初のクックソニアが現われてから4000万年もたってようやく、『葉』と優れた『配管システム』および地面を深く掘る『根』が誕生したのである。
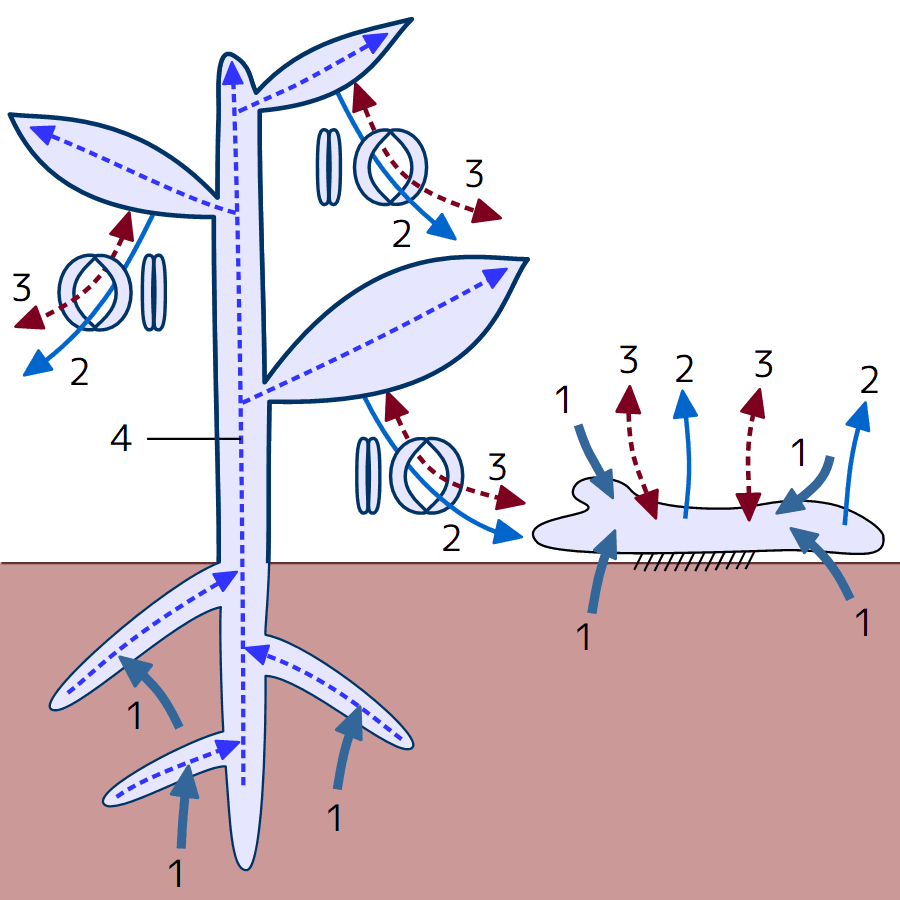
根を深く張ることには、植物にとって二つの利点がある。一つは、植物体の安定性を高めること、もう一つは、根を深く伸ばすことで土壌からの養分と水の取得量が増えることである。最初の陸上植物の根は浅かったが、葉の出現に伴い、根は進化を遂げて土壌の深部にまで達するようになった(注109)。
(注109)維管束植物(左)とコケ植物(右)の体内での吸水(1)、蒸発・蒸散(2)、ガス交換(3)、水の輸送(4)の働きを比較すると、図123のとおりになる。気孔が主にガス交換と蒸散(水の蒸発)を行う維管束植物の体表は、透水性・通気性が低いクチクラで覆われている。気孔の分布と孔辺細胞の変形による開閉機能によって、ガス交換の領域とタイミング・量が調節される。生命活動や蒸散に必要な水は、土壌から吸収され、導管を通って根から茎、さらには葉へと移動する。蒸散は、導管/仮導管に水が上昇する原動力となる。根は、土壌中の広範囲から水分と養分を集める能力を持ち、特定の菌類が生育する場(菌根)を提供することで養分の吸収力を高める。維管束植物では、吸水とガス交換の場所が分離され、吸水は根で、ガス交換は気孔で主に行われている。この気孔、根、そして両者を繋ぐ維管束の組み合わせが、コケ植物より大きく高い植物体を可能にした。
デボン紀の始まり頃には、すでに1メートル近くまで根を伸ばせる植物の存在が確認されている。また、陸上植物の個体数の増加に伴い、枯死した植物によって土壌中の有機物質が増加した。さらに、根が深く伸びるにつれて、その下の岩石は物理的・化学的風化作用を大きく受け、これが大気の組成や地球の気温に重大な影響を及ぼした。
これまで述べてきたように、二酸化炭素を大気中から除去する最も重要なプロセスは、『ケイ酸塩岩石の風化』である。これには主に、『花崗岩』や花崗岩に化学的に類似したケイ素を豊富に含む堆積岩や変成岩が関与する。ケイ酸塩岩石が風化すると、化学反応を通じて大気中の二酸化炭素が除去される(注110)。
(注110)ケイ酸塩岩石の化学的風化作用の一例を上げると、貫入火成岩体(注111)の周囲の接触変成帯に存在する「珪灰石(ウラストナイト)CaSiO3」は化学的風化作用
CaSiO3+CO2 → CaCO3+ SiO2
により大気中の二酸化炭素を固定する。
(注111)マグマが地表に噴出せずに地下で固結すると形成される。その形状により、「岩脈」、「岩床」、「バソリス(底盤)」、「岩株」、「ラコリス(餅盤)」、「ロポリス(盆盤)」、「シル」などが存在し、それぞれの特性が岩石の種類に影響を与える。
森林が大地を覆い始めると、生物が風化を促進する現象がすぐに起きたと思われる。これはおよそ3億8000万年前から3億6000万年前の出来事であった。根がケイ酸塩岩石の深部にまで到達するにつれて、花崗岩や花崗岩に似た岩石は、森林が現れる前の時代と比較して風化速度が格段に速まる。その結果、大気中の二酸化炭素濃度は急速に下降した。
二酸化炭素濃度が下がると、大陸には氷が現われる。初めは高緯度地方だけだが、次第に低緯度の地域にも拡大する。
進化の力は高木を優先し、木が高くなれば根も深くなる。植物の高さはますます伸び、根はさらに長くなり、地球はどんどん寒冷化していった。
- 陸上植物が登場してその根が深さを増していったことが、地球史の中でも最長規模となる『石炭紀の氷河期』をもたらした。
しかし、この氷河期が訪れるまで、世界は暖かく、樹木が豊富に生えていた。二酸化炭素の濃度も植物にとって理想的なレベルだったと考えられる。
- 大陸が維管束植物の緑で覆われるようになったことは、顧客が存在しないにも関わらず在庫が豊富な巨大な食料品店に例えられる。店内に入れば、無制限に食事ができたのである。
最初の陸上動物
陸上への適応を試みる生物にとって、一つの重大な問題は水分の保持であった。全ての生細胞は内部に液体を保有しており、水中生活においては脱水のリスクは存在しない。しかし、陸上での生活では、体内の水分を保つための強固な保護膜が必須となった。
- 難しいのは、体表の乾燥を減らそうと思うと、表面の膜を通した呼吸ができなくなることで、厄介な難題に挑むか、深く青い海を選ぶかの板挟みである。
外皮をコーティングすれば乾燥を防ぐメリットがある一方で、同時に窒息のリスクも増大する。体表面に呼吸器官を発達させて、酸素が体内に浸透するようにするという選択肢も存在するが、これは脱水リスクを高める結果となった。陸を征服したければこのハードルを乗り越えねばならず、その難しさゆえに、動物にせよ植物にせよ原生動物にせよ、水から陸への適応に成功した生物群は限られていた。
現存する海洋動物の中でも最大級でとりわけ重要な門は、とうとう最後までそれを成し遂げられなかったわけである。たとえば「海綿動物」、「刺胞動物」、「腕足動物」、「外肛動物」、「棘皮動物」には陸上のものがまったく存在しない。
- 最初の陸上生物の化石として確認されているのは小型の「節足動物」であり、現代のクモ、サソリ、ダニ、ワラジムシや原始的な昆虫に姿が似ている。先に進出したグループは特定できていない。このうちどのグループが先陣を切ったのかはわからない。
既知の化石は全て古い堆積物から発見されており、最初に進出した生物の存在は長くは続かなかったと推測される。最初の陸上生物を特定するためには化石記録に依存せざるを得ないが、小型の陸生節足動物の化石は不確かな情報源として知られている。それぞれのグループの外骨格は非常に石化しにくいため、化石として残ることが稀である。それでも、
シルル紀後期からデボン紀初期(約4億年前)にかけての陸上植物の出現は、陸地への最初の進出を促進した。『節足動物』の複数の系統が独自に空気呼吸システムを発達させたことは明らかである。
現代のサソリやクモの呼吸器系を観察することで、海洋生物から陸上生物への移行が可能となった理由の一部が明らかとなる。この重大な転換を達成する上で、呼吸器官の存在が極めて重要であった。最初の陸上節足動物の肺は過渡的な構造を持ち、後の種の肺の効率性とは大きく異なる可能性が高い。
しかし、
大気中の酸素濃度が高ければ、非常に小型の陸上生物は体壁を通じて酸素が浸透する可能性があり、また、原始的な肺構造でも酸素を取り込むことができた。そして事実上、最初の陸上生物は全て小型であった。陸上への進出を果たした動物門には、「節足動物」だけでなく、「軟体動物」、「環形動物」、「脊索動物」(そして非常に小型の線虫のような生物)も含まれていた。しかしながら、進化の過程で成功を収めたのは「節足動物」であった。
全体を覆う外骨格は既に乾燥から保護する役割を果たしていた。しかし、呼吸の問題は未だ解決しなければならなかった。節足動物の高次分類群が初めて化石記録に現れたのは酸素濃度が低かったカンブリア紀であった。その中で確実に生存を維持するために、節足動物はほぼ全ての体節の外骨格に大きな鰓を発達させる必要があった。しかしながら、そのような外鰓(がいさい)は空気中では機能しない。したがって、
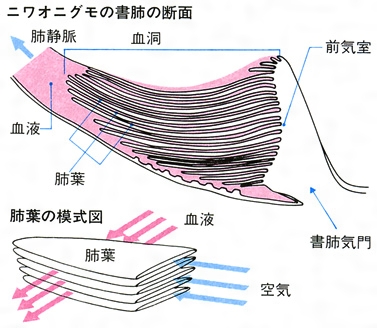
初めて陸上に適応した節足動物やクモ、サソリは、『書肺』と呼ばれる新たな呼吸器を発達させた。その肺の内部には多数の折れ曲がった構造が存在し、その姿が書物のように見えることからその名がついた。
- そのページの間には血液が流れ、甲皮に開いた多数の孔を通じて空気が書肺に入ってきた。
- 能動的に「吸い込む」わけではないので、受動的な肺といえる。この性質を持つことから、書肺が適切に機能するためにはある程度の酸素濃度が必要である。
広く知られているように、非常に小型のクモは、風により高い場所へと飛ばされ、「空中プランクトン」と呼ばれる存在がいる。これを考慮すると、低酸素環境でもクモは十分な酸素を取り込むことが可能と思われる。しかし、この種類のクモは非常に小型であるため、体壁を通じて少量の酸素が受動的に浸透するだけでも、呼吸には大きな影響を与えないかもしれない。一方、より大型のクモは依然として書肺に依存して呼吸している。
動物が初めて陸地に上陸した時期は明確には定まっていない。最古のサソリやクモは小型で、化石化しにくかったという事情がその一因である。現代のサソリはクモより石化しやすい性質を持つため、自然と化石として残りやすくなる。
- 陸上動物の化石断片が確認できる最も古い岩石は、ウェールズ地方に存在する約4億2000万年前の『シルル紀末期』に近いものである。
- その時代には、酸素がすでに非常に高い濃度に達しており、その時点で過去最高レベルとなっていた。
- 初期の化石は数がきわめて少なく、多様性も乏しい。それでも、その正体は特定されており、ほとんどの断片は『ヤスデ類』のものと見られている。
有名なスコットランドの『ライニー・チャート(4億1000万年前)』(注112)からは、格段に多様な生物群が見つかっている。この堆積層にはごく初期の植物のほか、小型の節足動物の化石も含まれている。ほとんどの節足動物は現代のダニやトビムシに類似している。どちらも植物の破片やくずを食べることから考えて、丈の低い原始的な植物を中心とする新しい陸上生物群の暮らしにうまく適応していたに違いない。ダニはクモ類の一部で、トビムシは昆虫である。現在、昆虫は地球最大の動物群であり、その中でも最も古いとされるのがトビムシである。
(注112)チャート(chert)は堆積岩の一種。主成分は二酸化ケイ素(SiO2、石英)で、この成分を持つ放散虫・海綿動物などの動物の殻や骨片(微化石)が海底に堆積してできた岩石である。断面をルーペで見ると「放散虫」の殻が点状に見えるものもある。非常に硬い岩石で、層状をなすことが多い。なお、「ライニー」とは、イギリス北部スコットランドの地名である。
古昆虫学者によれば、昆虫の数は約3億3000万年前の石炭紀前期の終わりまで少なく、陸上動物の中では重要性が低い存在であった。
当時、酸素濃度は現代並みになっており、さらに記録的レベルへと上昇していた。それがピークに達したのは約3億3000万年前の石炭紀後期の後半であった。
昆虫が飛行能力を獲得するのにもまた時間がかかった。紛れもなく飛んでいたと見られる昆虫が化石記録に広く現われるのは、3億3000万年前ほど前の地層からである。この新たな段階に入るとまもなく、途方もない進化の波が昆虫に押し寄せ、おもに飛行するタイプの新種が続々と誕生した。
これは適応放散の典型例である。新しい形態を獲得したことで、新たな生態的地位を占めることが可能になった。また、大気中の酸素濃度が高かったことがその放散を促進し、後押ししたことは間違いない。
最初に陸地に上陸したのは昆虫ではなく、その栄誉はサソリに与えられている。約4億3000万年前のシルル紀中期に、鰓を持つ原始的なサソリが、それまでの生息地であった淡水湖沼から陸地に上がったと考えられている。これらのサソリはおそらく、岸辺に打ち上げられた魚の死骸などを探していたと思われる。鰓は湿った状態で、大きな表面積を持つため、ある程度の呼吸が可能であったとされる。機能的な肺は存在せず、鰓が代用と して使われていたと推測される。
現時点で判明している事柄を時系列に沿って整理すると、
- サソリの上陸は約4億3000万年前であったが、生殖や呼吸に関しては完全に水から離れられていなかった可能性がある。
- その後、4億2000万年前にはヤスデが、4億1000万年前には昆虫が上陸した。
- しかし、多様な昆虫が出現するまでには、さらに3億3000万年前まで時間が経過した。
- これらの事象の流れと、大気中酸素濃度の推移との間に何らかの関連性があるのだろうか。
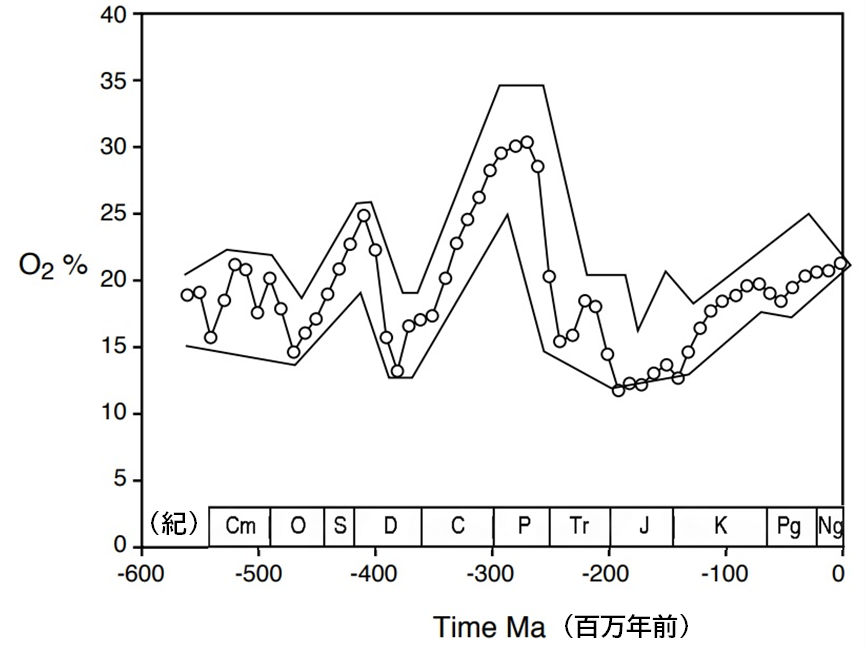
当時の酸素濃度に関する最新の推定値(<物語り その10>図114:右に再掲)によれば、高酸素の一つのピークはおよそ4億1000万年前に訪れた。その後酸素濃度は急落し、非常に低いレベル(12パーセント)となったが、デボン紀(Devonian period)の終わりに再び上昇し始め、ペルム紀(Permian period)の時期には史上最高レベルの30パーセントに達した。ライニー・チャートは、デボン紀における酸素濃度の最大期に堆積したもので、最初の生物群集である昆虫とクモ形類の化石を産出している。
- 昆虫の多様性を研究する古昆虫学者によれば、当時の化石記録では昆虫の数は非常に少なく、石炭紀(Carboniferous period)前期から後期(3億3000万年前~3億1000万年前)にかけて酸素濃度が20パーセント近くになった頃から、飛行昆虫の多様化が見られるようになった。
- 様々な脊椎動物が陸上に進出できた背景には、オルドビス紀(Ordovician period)からシルル紀(Silurian period)にかけての大気中の酸素濃度の上昇があったと考えられる。この酸素濃度の上昇がなければ、陸上への進出の経緯や動物の種類は、大きく異なっていた可能性がある。その極端な事例として、動物が全く水中から出てこなかった可能性もある。
- 陸上に進出したとはいえ、その時期の陸上動物の数は非常に少なかったと推察される。その理由の一つとして、大気中の酸素濃度が低下し始めたことが考えられる。

陸地で新たな生活を開始したのは、節足動物だけではない。軟体動物でもある『腹足類』(注113)も、進化の大きな飛躍を遂げて陸に上がった。しかし、この移行がなされたのは石炭紀後期に入ってからであり、したがって、第二の波に相当する。当時は、第一の波が起きた時代よりもさらに酸素濃度が高かった。軟体動物とほぼ時を同じくして、カブトガニも上陸している。
しかし、これらは生命史にとっては些細な存在であり、最も重要な動物群と目されるのは、私たちが属する『脊椎動物』である。それでもなお、このグループの中でも両生類が海から陸に進出したのは、長い進化の歴史の末のことであった。その詳細を検討する前に、一般的に『魚の時代』と呼ばれてきた『デボン紀』の状況を考察することが重要である。
(注113)腹足類は軟体動物門の一綱で、巻き貝を中心とするグループである。通常、巻いた貝殻の中に軟体が入り、体の腹面が全部足裏になっているのでこの名がある。殻は殻頂から殻口へしだいに太くなり、左右いずれかに巻いているが、多くが右巻きである。殻の蓋は足の背後についており、軟体を殻内に入れたとき殻口を閉じる役目をしている。軟体は普通、頭部に1対の触角と目をもつが、目を欠くものもある。口には歯舌がある。足は広くて平らな足裏になり、這うのに適していて、肛門や生殖孔は外套腔に開くようになっている。内臓は殻内に収まる。
ゴーゴー累層の魚
オーストラリアに位置する『ゴーゴー累層(注114)』という面白い名前の地層、ここの団塊の内部には今までに見つかった中でもとりわけ重要な化石が眠っていた。
(注114)「累層」とは、地層を岩相によって区分するときの基本になる単位層。同一の環境またはある環境の一定の繰り返しのもとで堆積した一連の地層で、上下の層とはっきり区別できるもの
この地層から発見された化石からは、私たちの祖先である「両生類」がどのような基盤の上に誕生したかが明らかになる。両生類の陸上進出について理解するには、まずデボン紀にどれだけ多様で複雑な魚の世界が広がっていたかを知らなくてはならない。
近年、オーストラリア・アデレードにあるフリンダーズ大学の古生物学教授ジョン・ロングが、新しい高解像度のスキャン技術を用いて画期的な発見をした。その発見からは、現存する魚すべての祖先についてや、人間のDNAに刻まれた太古の血統が見えてくる。
ロングの業績からわかるのは、デボン紀における魚類の進化、形態、多様性、および生態系が、教科書に書かれているよりはるかに複雑だったということである。これまで魚類は伝統的に四つのグループに分けられてきた。すなわち、『無顎類』(現存するのはメクラウナギとヤツメウナギ)、『軟骨魚類』(サメなど)、最も種類の多い『硬骨魚類』、そして今は完全に絶滅した『板皮類』(初めて顎を持った魚)である。

どれも長らく考えられていたほど単純なものではなかったことが、様々な点について判明してきた。ゴーゴー累層の発掘調査でロングは数々の発見を成し遂げており、
- 最古の硬骨魚である『ゴゴナスス』の完全な頭骨もその一つである。骨を調べたところ、頭頂部に大きな噴気孔をもつことが明らかにされた。それまでは魚類にも存在することが知られていなかったものである。
- さらには、他の種類の魚も予想以上に多様であり、新種の『肺魚』(最終的に陸に上がったものときわめて近縁)や「節頸類」と呼ばれる特異な魚が存在したことも明らかにした。
だが最も驚くべきは、「体内に胎児を宿した魚」の発見である。これは体内受精による生殖が初めて実証された事例であり、脊椎動物の胎生を示す最古の証拠となる。ロングの標本の一つでは、胎児とつながってへその緒のような構造も石化して残っており、その種の化石はほかには確認されていない。(注115)
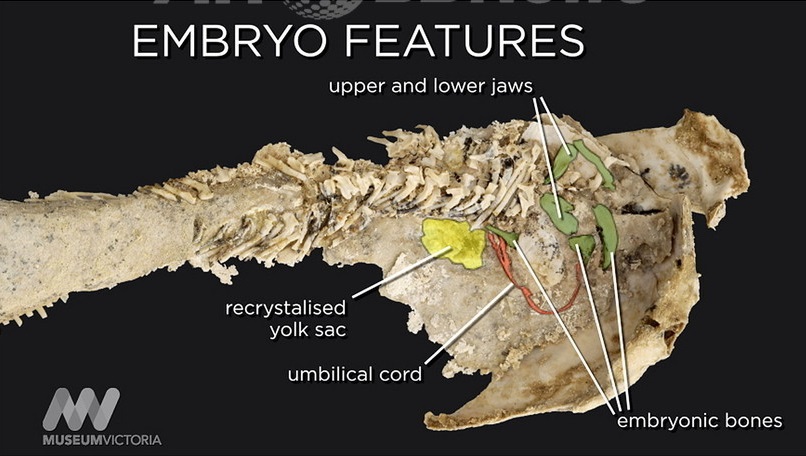
(注115)embryo:胚、embryonic bone:胎児骨、umbilical cord:臍帯、yolk sac:卵黄嚢
ロングは先進の技術を用いて、筋肉組織や神経細胞、微小毛細血管を三次元で見事に描き出した。これら全ては化石から初めて得られた情報である。しかしながら、
- 動物の陸上進出を理解する上で最も重要なのは、ロングが軟組織を明らかにしたことである。これにより、魚がどのように進化すれば子孫が歩行できるようになるのかを、新たな観点から考察することが可能となった。
陸生脊椎動物の進化
私たちが属する脊椎動物が、水生から陸生へと進化する過程は、最初の両生類の誕生から始まると考えられている。化石記録を通じて、この進化に関与した生物種とその時期は、ある程度特定されている。
最初の両生類の直接の祖先は、デボン紀の硬骨魚の一種である『扇鰭(せんき)類』(鰭:ひれ)(注116)であったと推定されている。当時のこの種は主に淡水に生息し、淡水生態系の主要な捕食者であった。これは注目すべき事実であり、陸地への進化の架け橋が海水ではなく淡水であったことを示唆している。「扇鰭類」には前適応が見られ、後に陸上での移動を可能にする四肢の発達につながる形質が存在していた。葉状で肉質のひれを持っていたのである。
(注116)硬骨魚は『条鰭類(じょうきるい)』と『肉鰭類(にくきるい)』の2系統に大きく分かれる。私たちが日常的に見る魚の大部分は「条鰭類」である。一方、胸びれの根元に骨と筋肉があるのが「肉鰭類」であり、その中の一種が『総鰭類』。総鰭類はさらに『扇鰭類(せんきるい)』と『シーラカンス類』に分けられる。
現存する『シーラカンス』は「生きた化石」と呼ばれ、最初の両生類がどのような生物から進化したのかを理解する上で非常に有用な参考例である。しかし、「肉鰭類」の別の下系統である『肺魚』も、進化の観点から重要である。肺魚は、移動能力ではなく、鰓から肺への移行、すなわち、呼吸器官の進化を考える上で非常に参考になる。どんなに優れた四肢を持っていたとしても、空気呼吸ができなければ陸上生活は不可能である。
- つまり肉鰭類には二つの重要な下系統がある。一つは「総鰭類」の系統で(シーラカンスはここに入る)、もう一つは「肺魚」の系統である。
扇鰭類から「両生類の祖先種」が分岐したのは、おおむね4億5000万年前、オルドビス紀からシルル紀への移行期と重なる時期であった。
- ただしこれは、両生類自体が誕生した時期ではなく、最終的に両生類へと進化する可能性を持つ「魚類の祖先種」が生まれた時期であったと考えられる。

古生物学者ロバート・キャロルによれば、
- 最初の両生類へと進化した最後の魚として最も有力な候補は『オステオレピス属』(注117)である。この魚類は、デボン紀初期から中期にかけての約4億年前まで生息していた。
(注117)オステオレピス属はデボン紀中期に生息していた絶滅した魚類の属である。体形は細長く、鱗は菱形、鰭は葉状であった。全長は約20cmで、南極地域に生息していたと推測されている。

アイルランドのバレンシア島では、非常に興味深い足跡が発見されている。これらの足跡から推定すると、最初に陸上で生活した両生類はこの頃に出現した可能性がある。この足跡は、四肢を持つ動物による最古のものとされ、年代は約4億年前と推定されている。これらの足跡は、太い尾を引きずりながら泥地を歩いた一匹の動物によってつけられたと考えられており、150個ほど確認されている。関連する骨格の化石は一切見つかっていないが、これらの足跡の発見は大きな議論を引き起こした。それは、これらの足跡が最初の四肢動物とされている骨よりも3200万年前のものだったからである。
- しかし、この足跡が形成された時代の酸素濃度は、現代のレベルに近いかそれを上回っていたと考えられている。これは重要な観察点である。なぜなら、陸生昆虫と陸生クモ形類の最初の化石が出現するのも同じ時代だからである。高い酸素濃度は、昆虫の陸上進出を助けただけでなく、最初の陸生脊椎動物の誕生も促進した可能性がある。
この足跡の形成時期についての疑問を少し和らげるような化石が2010年に発見された。別の場所で見つかった足跡の年代が3億9500万年前と特定されたのである。この足跡はポーランド南部の海洋性堆積物の中に保存されており、デボン紀中期のものである。いくつかの足跡には足指も確認できた。
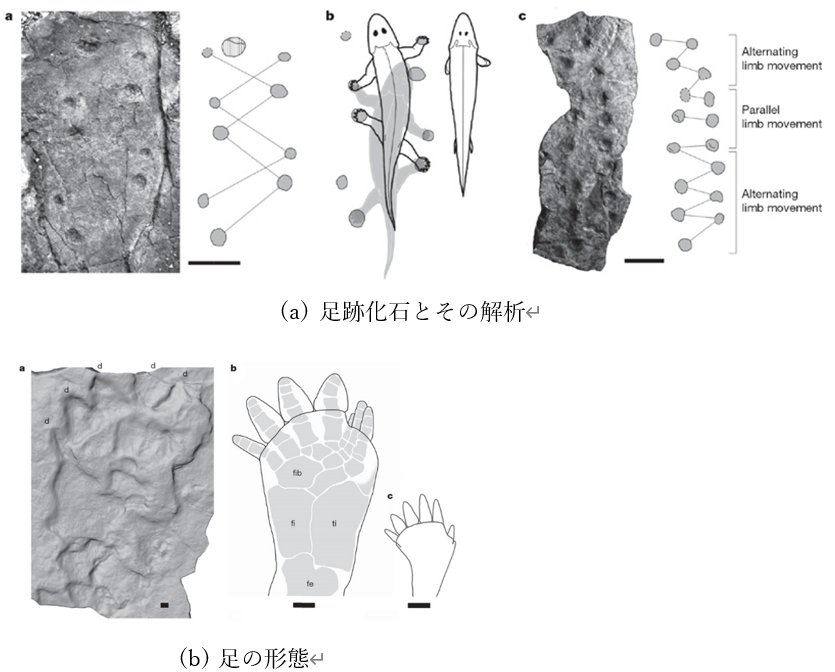
推定通りの年代だったとすれば、現時点で最古とされる四肢動物の体化石より1800万年古いということになる。
さらに興味深いことに、この足跡は、魚に近い四肢動物では達成不可能な特定の移動パターンを示唆している。これは、ティクターリクやその子孫と考えられるアカントステガとは異なる動物が存在していたことを示唆している。この足跡を残した動物は当時としては大型で、体長は240センチを超えていた可能性がある。干潟に生息し、打ち上げられた海洋生物や各種の陸生節足動物(クモやサソリを含む)を食べていたと推測されている。

現状では、四肢動物の骨の化石として最も古いものは3億6000万年前の岩石中から見つかったものである。これに基づき、
- 魚類から両生類への進化は、おおよそ4億年前から3億6000万年前までの期間に起こったと推定される。
- この間、特筆すべき特徴として、図114に示されるように酸素濃度が急激に低下したことが挙げられる。ロバート・バーナーの曲線からも、この時期は大気中の酸素が極めて少なかったことが確認できる。しかし、実際の生物の移行はそれよりも早い時期に起こったと考えられる。酸素濃度が低下し始めた時点では、依然としてデボン紀の高酸素濃度のピークに近い状態であったと考えられる。
両生類の誕生は、生物進化の観点から極めて重要な事象であるにもかかわらず、それを理解するための手がかりとなる化石は非常に少ない。その中でも、四肢動物の化石が最も多く見つかる場所はグリーンランドである。多くの文献では、イクチオステガ属が最初の両生類であるとされているが、
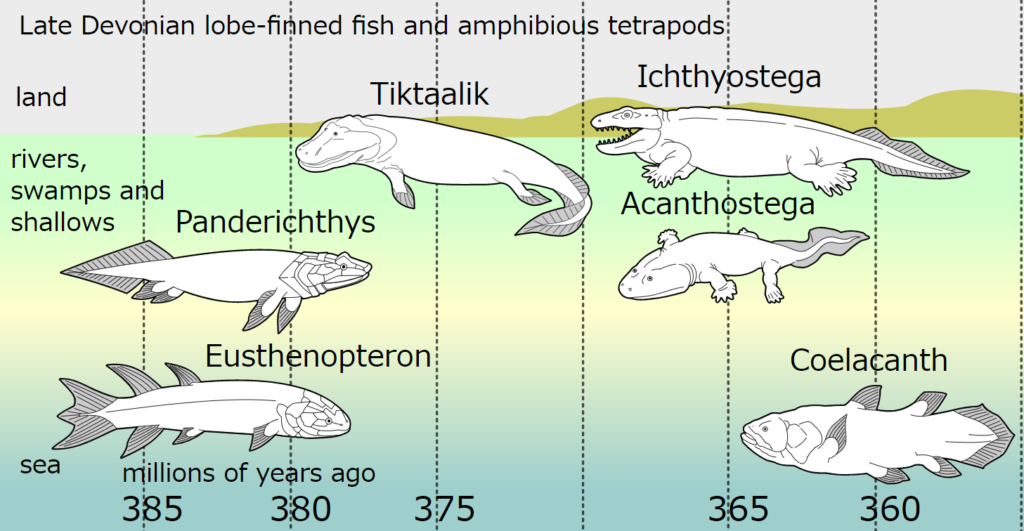

- 実際には、『ヴェンタステガ』属が最も古く、約3億6300万年前に出現したという事実がある。
- その後、数百万年を経てヴェンタステガが分布を広げた頃に、『イクチオステガ』、『アカントステガ』、『ヒネルペルトン』といった新たな属が誕生した。
イクチオステガは最も有名だったが、ティクターリクの発見以降はその名声をさらわれた感がある。しかしながら、ティクターリクが依然として魚である一方で、イクチオステガは両生類であるという事実を明確に理解することが重要である。イクチオステガの骨格は1930年代に初めて発掘されたが、それは断片的なものにすぎなかった。その完全な復元には1950年代までを要した。

イクチオステガは発達した脚を持ちながらも、魚のような尾を保持していた。その後の研究により、この3億6300万年前の生物が陸地を歩行する能力を持っていたとは考えにくいことが明らかになった。新たに分析された足首と足先のデータから、イクチオステガは水中の浮力がなければ体を支えることが困難であったと考えられる。
イクチオステガやその他の原始的な四肢動物が見つかったグリーンランドの地層は、デボン紀後期の大量絶滅直後のものである。この大量絶滅の原因は、間違いなく酸素濃度の低下であり(図114参照)、結果的に海は広範囲にわたって酸欠状態となった。イクチオステガとその仲間の登場は、この大量絶滅が引き金となった可能性がある。生物進化の新たな局面は、大量絶滅後に空いた生態的地位を埋めるために生まれるケースが多いからである。しかしながら、イクチオステガの繁栄も長くは続かず、登場からわずか数百万年後には姿を消した。
デボン紀後期に出現したイクチオステガとその同類からは、重要な疑問が生じる。すなわち、
- もしそれらが本当に最初の陸生脊椎動物であったのであれば、なぜその後、子孫が適応放散(起源を同一にする生物群が、種々の異なる環境に適応する形態的な分化を起こし、多くの系統に分かれる現象)を遂げることができなかったのか
である。
- 実際には、新たな両生類が出現するまでに長い空白期間が存在する。この空白期間は長年、古生物学者たちを悩ませてきた。そしてこの期間は『ローマーの空白』と呼ばれるようになった。
- 20世紀前半に活躍した古生物学者アルフレッド・ローマーの名にちなんだこの用語は、脊椎動物の上陸の第一波と第二波の間に存在する空白期間を指す。実際、両生類の適応放散は3億4000万年前から3億3000万年前まで起こらず、ローマーの空白は少なくとも3000万年にわたったと考えられる。
両生類が存在しないかに見えた空白期間を埋めたのが、イギリスの古生物学者ジェニファー・クラークである。2003年にクラークが博物館の古いコレクションを調査していた際、完全な水生魚とされていた化石に四肢と五本指が存在することに気づいた。しかもその骨格は、陸上生活に適した構造を有していた。
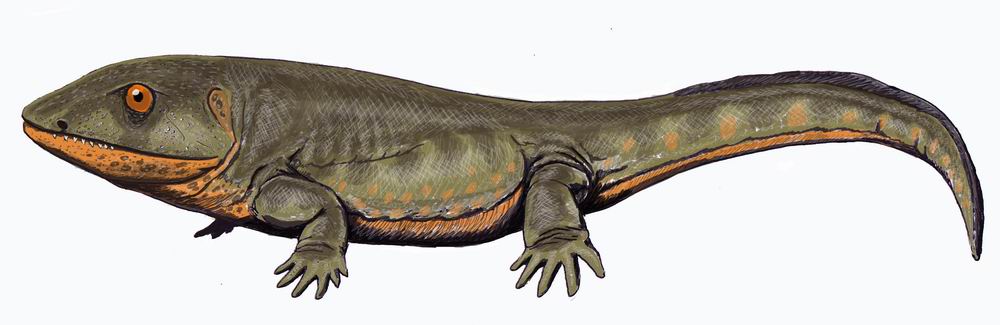
新たに『ペデルペス』(注118)と名付けられたこの化石は、ティクターリクよりも遙かに後の時期に生息していたことが明らかになった。それどころか、「ペデルペス」が真の両生類の先駆けである可能性があり、その存在期間(3億5400万年前~3億4400万年前)はローマーの空白期間と一致する。しかし、陸上で必要な脚がローマーの空白期間に発達していたことはわかっても、空気呼吸が可能であったか、また水から出られたのかどうかは依然として不明である。
(注118)ペデルペスは、約3億5,000万年前(石炭紀前期)のスコットランドに生息していた絶滅両生類である。化石では尾部が失われていたが、推定全長1m程度の湿地や浅瀬に住む生物だったと考えられている。頭骨は幅が狭く高さがあり、頑丈な歯を持っていた。「イクチオステガ」や「アカンソステガ」のようなデボン紀に生息していたまだ陸上に完全に適応できていない最古の両生類と、石炭紀後期の『エリオプス』(注119)のような陸上に適応した両生類を繋ぐ化石記録は長い間知られていなかった。この間の約2,000万年の期間、『ローマーの空白』と言われていた空白のさなかに生息していたミッシングリンクが『ペデルペス』である。1971年に発見されて以来魚類のものだと思われていた化石が四肢動物であることを2002年に指摘したのがジェニファー・クラークである。
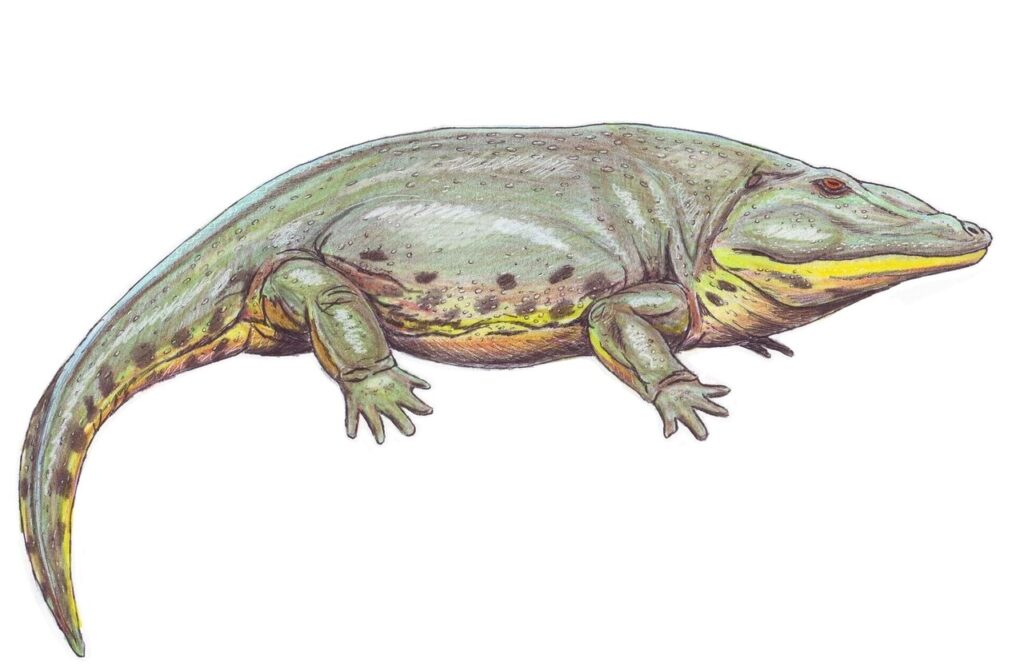
(注119)古生代ペルム紀前期の北米に生息していた半水生の絶滅両生類。学名は「引き伸ばされた顔」の意。この時期で最も有名な両生類であり、四肢動物の陸上への適応の例としてよく上げられる。全長2メートルほどで、体重は推定90キログラム。学名の示すように、頭部は非常に大きく50センチメートルを超え、幅広くやや扁平で、口には顎骨の縁のみならず口蓋部にも頑丈な歯が並んでいた。物を噛み砕く機能は無く、ワニ類のように逃げられないよう歯で押さえつけた獲物を頭を後方に振り上げることで丸呑みしていたと思われる。
イクチオステガやペデルペスのような水生の四肢動物が陸生へと移行するには、ティクターリク級の魚の構造を備えたうえで、
- 手首、足首、背骨、その他の中軸骨格を変化させる必要がある。さもないと陸上での呼吸と移動ができない。
- 胸郭は肺を収めるうえで重要である。
- また、重い体は水中では浮力に頼ることができたが、空気中で支えるためには肩帯や骨盤領域、さらにはそれを結合する軟組織を大幅に変えなくてはならない。
これらの変化をすべて成し遂げた最初の生物こそが、最古の陸生両生類と呼ぶにふさわしい。
もっとも、空気呼吸ができる呼吸器官と、重い体で陸上を移動できる四肢を発達させれば、すぐにでも両生類が著しく多様化するかに思えるが、実際には3億4000万年前~3億3000万年前になるまでそれは起きなかった。
- しかし、ついに適応放散が始まると目覚ましく分布域を広げ、石炭紀前期の終わり頃(約3億1800万年前)にはおびただしい数の両生類が世界中に生息するまでになった。
今ある証拠から判断すると、両生類の構造(つまり陸で暮らせる魚の構造)は一度で発達したのではなく、二度ないし三度にわたった可能性がある。
- 最初は約4億年前で、アイルランド・バレンシア島の足跡や「ティクターリク」化石の発見によって裏づけられる。
- 二度目は約3億6000万年前(「イクチオステガ」)であり、
- 最後が約3億5000万年前(「ペデルペス」)である。
- 長らく最初の陸生脊椎動物とみなされていた「イクチオステガ」は、当初考えられていた以上に魚に近かったと思われる。鰓を失っていても、それだけでは完全に陸上で暮らせた証拠にならない。
現に、今いる魚の100種類以上が鰓呼吸と併せて、ある種の空気呼吸を行っていることが確認されている。このうち68種については空気呼吸が個別に進化しており、これがいかに発生しやすい適応かを物語っている。
イクチオステガは、ほかの四肢動物の系統とつながってすらいなかった可能性もあり、最終的には完全水生の暮らしに戻ることになる。原始的な肺しか備えていなかったところに、デボン紀後期の酸素濃度低下が重なって陸を追われたのである。
初期の両生類は淡水性であったというのが長年の定説であったが、実際にそうであったのかどうかは、生命史上の大きな謎の一つであった。上陸は淡水を介して行われたのか、それとも海水から陸へと直接進化した生物も存在したのか。最新の研究によれば、初 期の総鰭類と肺魚の多くは海水性であったことが明らかになっている。カナダの古生物学者ミシェル・ローランも同様の指摘をしている。初期の両生類化石を産出する代表的な石炭紀地層の一部は、長らく淡水の堆積物と考えられていたが、実際には海または海に近い環境(潮間帯や潟など)の堆積物であったのではないかと推察されている。それでも、有名なティクターリクを含む、イクチオステガやアカントステガなどの初期の両生類が淡水性であったと解釈できるのは同じくらい確かである。
その結果として、これらの初期の両生類と両生類に近い生物は多種多様な環境で生息していた可能性が高いとされている。それは海水、淡水、そして陸上も含む。
そう考えると興味深い問題が浮かび上がる。現代の両生類は海水には耐えられない。その皮膚は、水に浸っているときに酸素を取り込むことができるが、海水には対応できない。これはおそらく、進化の歴史のかなり後になって獲得した形質に違いない。
まとめると、動物の上陸は二度(4億年前と3億5000万年前)にわたって起こり、いずれも酸素濃度が高かった時期と一致する。その間には、『デボン紀後期の大量絶滅』や『ローマーの空白』([補注]約3億6000万年前から約3億4500万年前までの約1500万年間、デボン紀末の原始的な森林と多種多様な魚類が、より派生的な前期石炭紀の水中および陸上の生物集団と分断されている)と呼ばれる時期が存在し、陸上で生活する動物はほとんど存在しなかった。したがって、「ローマーの空白」という概念には、「脊椎動物」だけでなく「節足動物」や「脊索動物」(注120)も含めるべきである。この空白期間は石炭紀に終焉を迎え、その時代には大気中の酸素濃度が大幅に上昇し、石炭紀末からペルム紀にかけてピークに達し、約32~35パーセントとなった。この高濃度が地球の歴史でほかに類を見ない特徴的な時代、すなわち『巨大生物の時代』を生み出したのである。
(注120)原索動物門(一時期あるいは終生「脊索」を持ち、「脊椎」は持たない動物)と脊椎動物門とをあわせた動物群に対する名称。脊索動物は、発生の初期に脊索(柔軟な棒状体で、中胚葉由来の細胞から構成され、胚の初期の軸を規定)を生じるのが第一の特徴で、脊索が終生続くものと、椎骨に置き換えられて縮小するものとがある。
図表
図117 ティクターリクの想像図
Tiktaalik roseae By: Michelle Raabe and Casey Wojtowicz
(http://bioweb.uwlax.edu/bio203/f2013/raabe_mic2/index.htm)
図118 緑藻の例:糸状性のアオミドロ
フリー百科事典『ウィキペディア(Wikipedia)』「緑藻」
(https://ja.wikipedia.org/wiki/%E7%B7%91%E8%97%BB)
図119 初期の陸上植物
コトバンク、日本大百科全書(ニッポニカ)「化石」
(https://kotobank.jp/word/%E5%8C%96%E7%9F%B3-44750#E6.97.A5.E6.9C.AC.E5.A4.A7.E7.99.BE.E7.A7.91.E5.85.A8.E6.9B.B8.28.E3.83.8B.E3.83.83.E3.83.9D.E3.83.8B.E3.82.AB.29)
図120 カタシャジクモ(硬車軸藻)
フリー百科事典『ウィキペディア(Wikipedia)』「カタシャジクモ」
(https://ja.wikipedia.org/wiki/%E3%82%AB%E3%82%BF%E3%82%B7%E3%83%A3%E3%82%B8%E3%82%AF%E3%83%A2)
図121 保護膜としてのクチクラ
産総研マガジン、研究成果記事一覧2013年「植物の表面を覆うクチクラ形成に重要な制御遺伝子
を発見」
(https://www.aist.go.jp/aist_j/press_release/pr2013/pr20130524/pr20130524.html)
図122 クックソニアの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「クックソニア」
(https://ja.wikipedia.org/wiki/%E3%82%AF%E3%83%83%E3%82%AF%E3%82%BD%E3%83%8B%E3%82%A2)
図123 維管束植物(左)とコケ植物(右)の吸水(1)・蒸発/蒸散(2)・ガス交換(3)・水輸送(4)の比較
福原のページ(植物形態学・生物画像集など)、Web site of FUKUHARA, T. (Fukuoka Univ. of
Education): Plant Morphology & Systematics、0-3.陸上植物を構成するグループ
(https://staff.fukuoka-edu.ac.jp/fukuhara/keitai/0-3.html)
図124 ニワオニグモの書肺の断面
コトバンク、日本大百科全書(ニッポニカ)「書肺」
(https://kotobank.jp/word/%E6%9B%B8%E8%82%BA-80559)
図125 腹足類の例
フリー百科事典『ウィキペディア(Wikipedia)』「腹足綱」
(https://ja.wikipedia.org/wiki/%E8%85%B9%E8%B6%B3%E7%B6%B1)
図126 ゴゴナススの頭部化石
奇跡の生物・古代魚の意外な共通点
(http://kiseki2seibutu.cocolog-nifty.com/blog/2010/11/post-a747.html)
図127 へその緒でつながった子を体内に持つ魚の化石
AFPBB News、環境・科学、「「胎児とへその緒」をもつ最古の魚の化石、オーストラリア
で発見」
(https://www.afpbb.com/articles/-/2397647?pid=2976502)
図128 硬骨魚類の系統樹
東京大学コレクションII、「動く大地とその生物、魚類」
(https://umdb.um.u-tokyo.ac.jp/DKankoub/Publish_db/1995collection2/tenji_gyorui_29.html)
図129 オステオレピスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「オステオレピス」
(https://ja.wikipedia.org/wiki/%E3%82%AA%E3%82%B9%E3%83%86%E3%82%AA%E3%83%AC%E3%83%94%E3%82%B9#:~:text=%E3%82%AA%E3%82%B9%E3%83%86%E3%82%AA%E3%83%AC%E3%83%94%E3%82%B9%20(Osteolepis)%20%E3%81%AF%E5%8F%A4%E7%94%9F%E4%BB%A3%E3%83%87%E3%83%9C%E3%83%B3,%E5%85%A8%E9%95%B7%E3%81%AF%E7%B4%8420cm%E3%80%82)
図130 バレンシア島の足跡化石
ナオコガイドのアイルランド日記
(http://naokoguide.com/blog-entry-793.html)
図131 デボン紀中期の足跡化石
Grzegorz Niedźwiedzki, Piotr Szrek, Katarzyna Narkiewicz, Marek Narkiewicz &
Per E. Ahlberg “Tetrapod trackways from the early Middle Devonian period of
Poland”Nature 463(2010),43–48
(https://www.nature.com/articles/nature08623)
図132 アカントステガの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「アカントステガ」
(https://ja.wikipedia.org/wiki/%E3%82%A2%E3%82%AB%E3%83%B3%E3%83%88%E3%82%B9%E3%83%86%E3%82%AC)
図133 デボン紀後期の総鰭魚類と水陸両生四肢動物
フリー百科事典『ウィキペディア(Wikipedia)』「四肢動物」(https://ja.wikipedia.org/wiki/%E5%9B%9B%E8%82%A2%E5%8B%95%E7%89%A9)
図134 ヴェンタステガの想像図
File:Ventastega life restoration white background.jpg
(https://en.wikipedia.org/wiki/File:Ventastega_life_restoration_white_background.jpg)
図135 イクチオステガの想像図
ウィキメディア・コモンズ
(https://commons.wikimedia.org/wiki/Main_Page?uselang=ja)File:Ichthyostega BW 2.jpg
図136 ペデルペスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「ペデルペス」
(https://ja.wikipedia.org/wiki/%E3%83%9A%E3%83%87%E3%83%AB%E3%83%9A%E3%82%B9)
図137 エリオプスの想像図
フリー百科事典『ウィキペディア(Wikipedia)』「エリオプス」
(https://ja.wikipedia.org/wiki/%E3%82%A8%E3%83%AA%E3%82%AA%E3%83%97%E3%82%B9#:~:text=%E3%82%A8%E3%83%AA%E3%82%AA%E3%83%97%E3%82%B9%EF%BC%88Eryops%EF%BC%89%E3%81%AF%E3%80%81%E5%8F%A4%E7%94%9F%E4%BB%A3,%E6%A8%A1%E5%BC%8F%E7%A8%AE%E3%81%AF%20Eryops%20megacephalus%E3%80%82)
図138 無脊椎動物から脊椎動物への進化
日本財団の助成により開設された「新世紀を拓く深海科学リーダーシッププログラム
(Hadal Environmental Science/Education Program)」
(https://www.aori.u-tokyo.ac.jp/project/hadeep/pd1.html)

{kind=link}