
第四部 生命の進化
第6章 動物の多様化と酸素濃度【5億 ~ 3億6000万年前】
『サンゴ礁』は進化が生み出したものとして非常に古い歴史を持ち、カンブリア爆発に続く生物多様性の増加と並行してその存在感を増してきた。
- 生物多様性の拡大であるカンブリア爆発は、はるかに大規模なオルドビス紀の多様化を導く進化の前段階であり、それを推進する役割を果たした。
- そして、種の数が急増する過程において、『サンゴ礁』が非常に重要な役割を果たしたのである。

最初の『礁』(生物がつくる三次元構造で、波によって流されないもの)の出現はカンブリア紀の最初期にまで遡る。最初の礁はサンゴによるものではなく、絶滅してしまった古代の海綿動物『古杯類』(注94)によるものだった。「サンゴ礁」の誕生はそれより少し遅く、オルドビス紀に入ってからである。そして、デボン紀が始まる頃には、その規模、多様性、分布域が大幅に広がっていた。その後も特徴的な生態系として常に存在し続けるが、ペルム紀末には他の多くの生物とともにその多くが絶滅することとなった。
(注94)海綿に類似した海生の着生動物で、盃のような形をした無脊椎動物であり、先カンブリア時代後期から古生代カンブリア紀前期に繁栄し、最初の「造礁生物」の1つとされている。海綿に類似した化石でサンゴと海綿の中間的な性質をもつ独立した種類とみなされ、分類学上は門に位置づけられる。昔から分類上の位置が論議され,石灰藻,サンゴ,海綿などとみなされてきた。円錐あるいは円筒形の石灰質骨格が,内・外壁の 2層からなって、各壁に大小多くの穴があいている。
古生代のサンゴ礁は、一見すると現代のそれと意外なほど似ている。礁の主役はサンゴであり、サンゴが礁の三次元構造を形成するレンガの役割を果たしている。それらは各種のモルタルで接合され、被覆状サンゴがセメントの役割を果たし、サンゴの頭や枝をつなげて巨大で複雑な石灰岩の基盤を築いている。しかし、より詳しく観察すると、4億年前のサンゴが現代のものとは外見も種類も異なることが明らかになる。サンゴの大きな頭の形は現代と近いようでいて、細部の形態が大きく異なるのである。


- 現代のサンゴ礁で最も一般的なのは「石サンゴ」だが、4億年前に同等の地位を占めていたのは『床板サンゴ』であった。床板サンゴは半球状に枝分かれし、その間には他の「造礁性」を持つレンガも存在していた。
- その多くは『層孔虫類』(注95)と呼ばれる奇妙な海綿状の生物で、炭酸塩を分泌していた。現在も存在しているが、大きさや多様性は古生代に比べて格段に小さい。
(注95)層孔虫類は海中に生息する微小な生物で、炭酸カルシウムを分泌し、殻を作る能力を持つ。現代でも生存しているが、その多様性や個体の大きさは古代のものと比べて小さい。これらの生物は地質時代の推定や古環境の復元に利用されることもある。
- この二種類の大型造礁生物のほかにも、別種のサンゴが点在していた。『四放サンゴ』と呼ばれるもので、群生を形成せず、牛の角のような形状を持つ。ただし、角の尖った部分、すなわち四放サンゴの炭酸カルシウムの骨格が土台と連結している。太い端は上方を指し、そこには幅広いイソギンチャクに似た生物が一固体固着している。
現代の石サンゴは、どれだけ大型で、どれだけ多数の『ポリプ』(触手をもつサンゴの構成単位)でできていようと、全体で一個の「個体」、少なくとも遺伝子レベルでは、と見なされる。これは4億年前の床板サンゴも同様である。時代を問わずどんな種類のサンゴも、イソギンチャクに似た微小なポリプが集まって形成する巨大な「コロニー」である。ポリプは、中央の小さな口を囲むようにして毒のある触手を生やしている。ただし、小型のイソギンチャク(単独生活するポリプ)が海岸の岩を覆い尽くす様子とは異なり、サンゴの個々のポリプは「共肉」(注96)と呼ばれる薄い組織で繋がっている。ポリプのコロニーはときに非常に大規模なものになるが、その遺伝子はすべて同一だ。それでいて、サンゴは一匹の動物とは言えない。その理由は、造礁サンゴが多種多様な植物を体内に共生させているからである。この共生植物は単細胞の「渦鞭毛藻類」(注97)と呼ばれ、ポリプ同士をつなぐ共肉の中やポリプ自体の体内に無数に存在する。この共生は両者に大きな利益をもたらす。微小な藻類は、生命の維持に必要な四つの要素、つまり光、二酸化炭素、栄養分(リン酸塩と硝酸塩)、そして様々な生物による捕食からの保護を受けることができる。
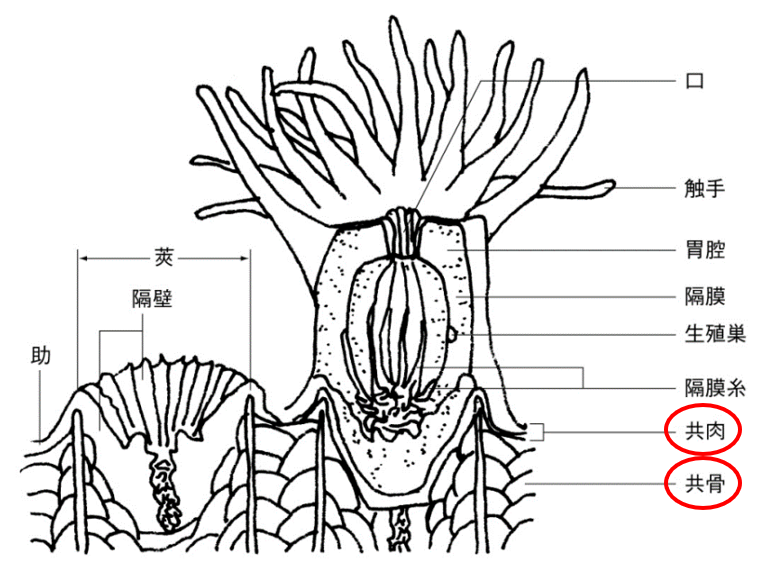
(注96)ポリプは「触手」で餌を捕らえ、「口」で食事や排泄を行い、「胃腔」で食べたものを消化する軟組織と、「莢」(さや)と呼ばれる骨格から成り立っている。この軟組織の構造はイソギンチャクと同じで、サンゴはいわば骨を持つイソギンチャクとも言える(図110)。サンゴは個々のポリプが「共骨」と「共肉」によってつながり、群体を形成している。群体は石灰質の骨格を形成しながら、ポリプが出芽・分裂を繰り返すことによって成長する。
(注97)2本の鞭毛を持つ単細胞生物で、海産プランクトンの重要な構成種であり、しばしば赤潮の原因となる。形態は多岐にわたるが、縦横2本の「鞭毛」を持つことが最大の特徴である。一本は鞭状のものを後方に引き、もう一本は羽状のものを横向きの溝に沿って巻き付ける。推進力の異なる2系統の鞭毛を備えるために、急停止や急発進、方向転換など、多様な遊泳行動が可能である。
オルドビス紀の多様化 - カンブリア爆発を土台に
カンブリア紀を終焉させた大量絶滅により、それまで大いに成功を収めていた多数の動物種が影響を受けた。生命の歴史の初期に登場した「三葉虫」や「腕足動物」、またバージェス頁岩に残された「アノマロカリス」のような風変わりな節足動物(<物語り その9>図96)も含まれる。この絶滅事象が発生したことは以前から認知されていたが、絶滅の規模は海洋生物種の半数以上が失われるほどではなかったと考えられている。
この絶滅が触媒となって、さらなる生物多様化の火種が点されたのではないかと推測される。適応性の低い生物種は絶滅し、新たな生物種と新たな機軸が登場する道が開かれた。
そして、生物種は全く新しい生活様式と生息地を発見することになった。カンブリア紀にはあまり利用されていなかった「汽水域」(河川水と海水が接触、混合する部分で、淡水域と海域の推移帯である。塩分が 0.5‰から 30‰までの範囲の水域を指す)や「淡水域」、また同じ海でもより深い海域や波打ち際に近い浅い海域への生物の進出が進んだ。動物は多くが依然として固着性で、一生の間に一箇所から移動せず、ますます栄養豊かになる海洋プランクトンを濾過摂食していた。それにもかかわらず、種の数もバイオマスも増加したのである。
オルドビス紀に入ると、カンブリア紀にはまだ見られなかった多種多様な動物が現れるようになった。これらの動物の大半はカンブリア紀末の大量絶滅の直後に出現した。その結果、動物相はカンブリア紀とは大幅に異なった構成になった。三葉虫は依然として生息していたが、カンブリア紀の海において最も一般的な生物であった三葉虫は、オルドビス紀においては殻を持つ他の動物によって、個体数及び種数ともに圧倒されるようになった。具体的には、腕足動物や多数の軟体動物が該当する。
最も顕著に進化したのは、全く新しい生活様式を進化させた群体性の動物であった。もちろん、群体を形成すること自体は、すでにより単純な体制の動物が行っていた。これらには様々な植物、微生物、原生動物などが含まれる。
しかし、オルドビス紀の群体性動物が特異であった点は、絶え間ない『多様化』を推進する原動力となったことであり、これこそがこの時代最大の特徴であった。特に注目すべき存在は『サンゴ』、『外肛動物』(注98)、そして新種の『海綿動物』であった。

(注98)外肛動物は小さな群体を形成して生活する動物で、サンゴに似た炭酸カルシウムなどの外壁からなる群体を作る。一般には『コケムシ』と呼ばれ、温帯から熱帯の海を好み、世界中に分布している。約8,000種が確認され、その数倍の種類の化石が知られている。群体は、山型、扇型、小枝型、栓抜き型など様々な形状を取る。外壁には小さな穴が無数に開いており、それぞれが「個虫」と呼ばれる個々の個体であり、口から肛門まで続く消化管からなる「真体腔」(注99)の構造も持っている。
(注99)「体腔」とは動物の外胚葉(体壁)と内胚葉(消化管)の間の空間であり、その中に内臓を収める。体腔の存在は動物群の系統や分類上重要である。脊椎動物では、体腔は中胚葉性の組織(腹膜など)で覆われている。また内臓の表面にも中胚葉性の細胞層があり、体腔は中胚葉の中に生じた空間である。ヒトの場合、多くの内臓は腹部の腔所に収まり、これを「腹膜腔」と呼ぶ。この腔所の上端は横隔膜で区切られているが、その上には肺と心臓があり、それぞれに区切られた腔所に収まっている。それらをそれぞれ「胸膜腔」、「囲心腔」といい、これら三つの体腔がヒトにおける体腔である。様々な動物群を比較すると、体腔の存在形態は多種多様である。例えば脊椎動物における腹膜腔、胸膜腔、囲心腔などが体腔であり、内胚葉由来の各器官が収まっている。このような前後三つの体腔を持つことは「三体腔性」と呼ばれる。体腔がない例や、空間が存在するが中胚葉の裏打ちがない場合を「偽体腔」と呼び、「真の体腔(真体腔)」と区別する。
この大規模な多様化の要因を探ると、「酸素」へと行き着く。
- 海水中の酸素濃度の上昇がもたらす真の影響は、この視点から捉えることで明らかになると思われる。そして、その解釈によって動物の多様化という問題の全体像が視えてくる。
- 言い換えれば、時間とともに動物の多様性が変動した主要な要因は「酸素濃度」であった。
地球上の動物の多様化が二部構成で進行したとすれば、オルドビス紀はその第二部に当たる。第一部はカンブリア爆発であり、どちらのケースも酸素濃度の上昇が原動力であった。オルドビス紀では、カンブリア紀と同様に、後の時代よりも速いペースで新種と新しい体制が現れた。これほど速く進んだ進化と革新は、生命史上初めて全世界に動物が満ち溢れた結果の一つである。
- カンブリア紀を一言で表現すれば、海が実験的な生物で満ちた時代であった。
- カンブリア紀以降の歴史を一言で述べるならば、生物間の生存競争により、適応度の低いものが無情に淘汰され、初期の原始的で非効率的なデザインが駆逐され、結果として生物多様性が急増した過程である。
- 進化は、優れた体制を探求し創出する手段となった。
生物多様性の歴史
ここで、「生物多様性の変遷」を検証してみよう。21世紀初頭にこの問題に注力したのは、ハーバード大学のチャールズ・マーシャル(現在はカリフォルニア大学バークレー校所属)と、当時カリフォルニア大学サンタバーバラ校にいたジョン・アルロイが指導する大規模な研究チームであった。このチームは、20世紀後半までのやり方が、単に図書館の科学文献に記載された過去の地質年代の種の数をまとめるというものだったのに対し、博物館の実際のコレクションを用いてより包括的なデータベースを構築した。するとほとんどの人にとって意外だったことに、長年の通説を覆す結果が得られたのである。
マーシャルとアルロイのチームの分析によれば、
- 古生代と中生代中期の生物多様性はほぼ同じであり、以前考えられていたような一貫した多様性の増加傾向は認められなかった。
- この結論の意味するところは大きい。何億年も前から、我々は生物多様性の安定した段階に入っている可能性があるのである。初期の生物の多様性がピークを迎えた後、その後はほぼ同じレベルを維持しているかもしれないし、すでに減少が始まっている可能性さえある。
確かに、新たな適応構造(例えば動植物の上陸を可能にするものなど)の出現により、地球の生物種に新たな種が追加されていたことは間違いない。それでも、古生代後期になると、種の数はほぼ頭打ちになっている可能性がある。つまり、
- カンブリア紀前期に生物が急激に多様化した後、種の数は急速に増加し、古生代を通じてその状態を維持した。しかし、ペルム紀末には激減した。
- その後は、全体として多様性が増加する傾向を示しながらも、「大量絶滅」と呼ばれる種の減少をもたらす短期的なイベントに度々遮られた。特に規模の大きかった五つの大量絶滅イベント(『ビッグファイブ』<物語り その9>)
- オルドビス紀末(O-S境界)
- デボン紀末(F-F境界)
- ペルム紀末(P-T境界)
- 三畳紀末(T-J境界)
- 白亜紀末(K-Pg境界)
が非常に重要な役割を果たした。大量絶滅が起きるたびに分類群は著しく失われるものの、その都度、種が形成されるペースが上がるため、絶滅以前のレベルどころか当初の多様性を上回る。
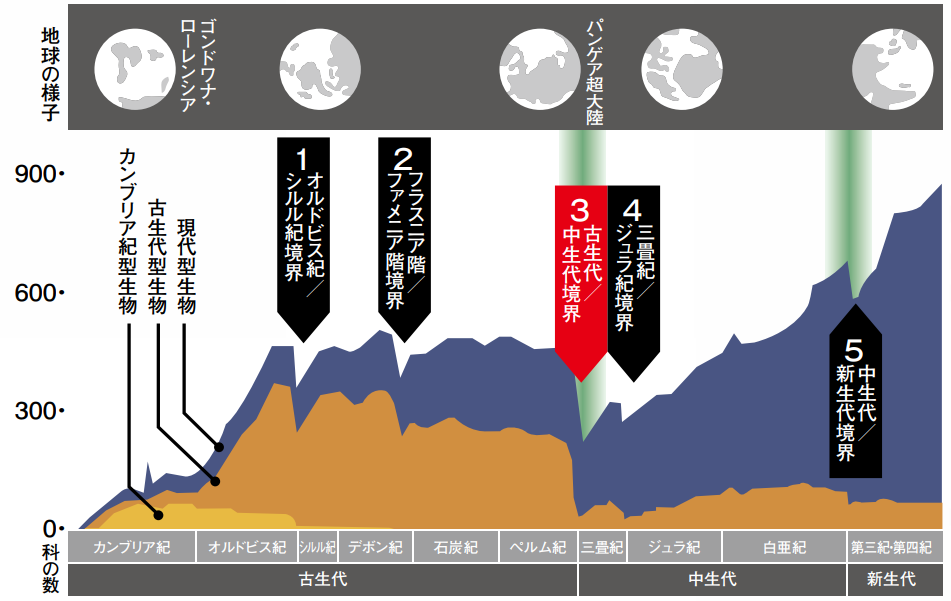
この歴史から見えてくるのは、顕生代を通じて観察される多様性のパターンが、多様化と絶滅を引き起こす数々の複雑な要因によって影響を受けているという事実である。多様性の増加につながる要素には、進化的な新機軸、新たな生息地への進出、新たな資源の出現などがある。一方、多様性の減少を引き起こす主要な要因には、気候変動、資源や生息地の減少、新たな生存競争や捕食、小惑星衝突のような外部イベントが含まれる。
特に、『酸素』が生物多様性とどのように関連しているかを示すデータが、2009年にイェール大学のロバート・バーナーによって提示された。
バーナーは、顕生代を通じての酸素濃度の変動(図114:エラーバーは予測モデルの誤差を示す)と、ジョン・アルロイのチームによる多様性の推移(図115:当時の最新データ)との間に重要な相似性を見つけ出した(図116参照)。
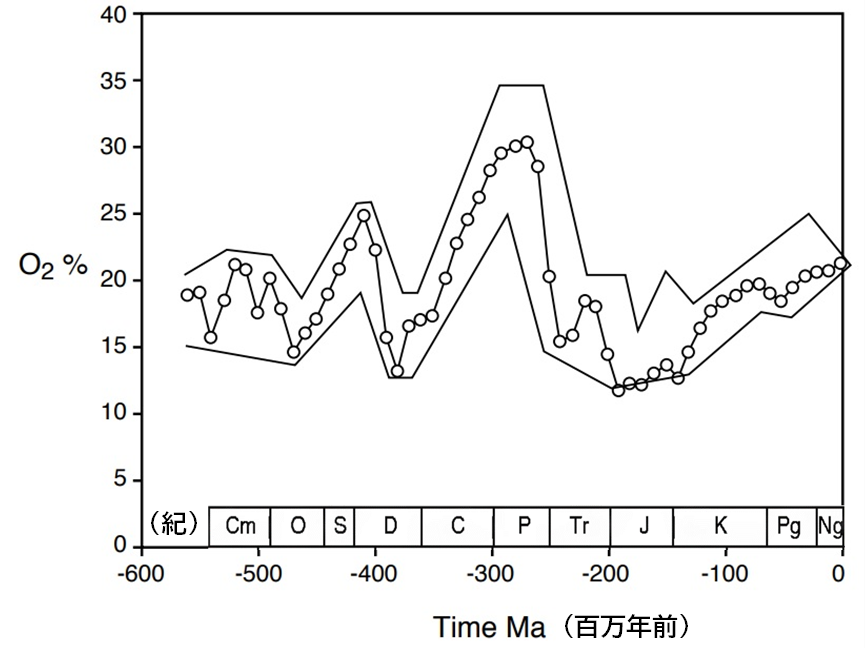
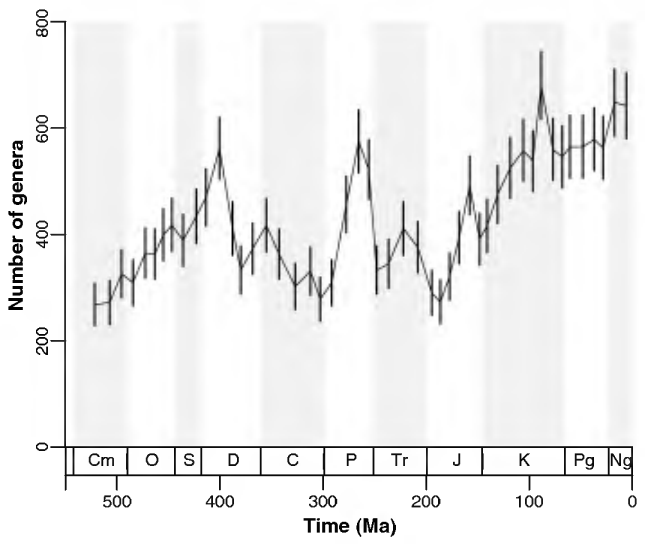
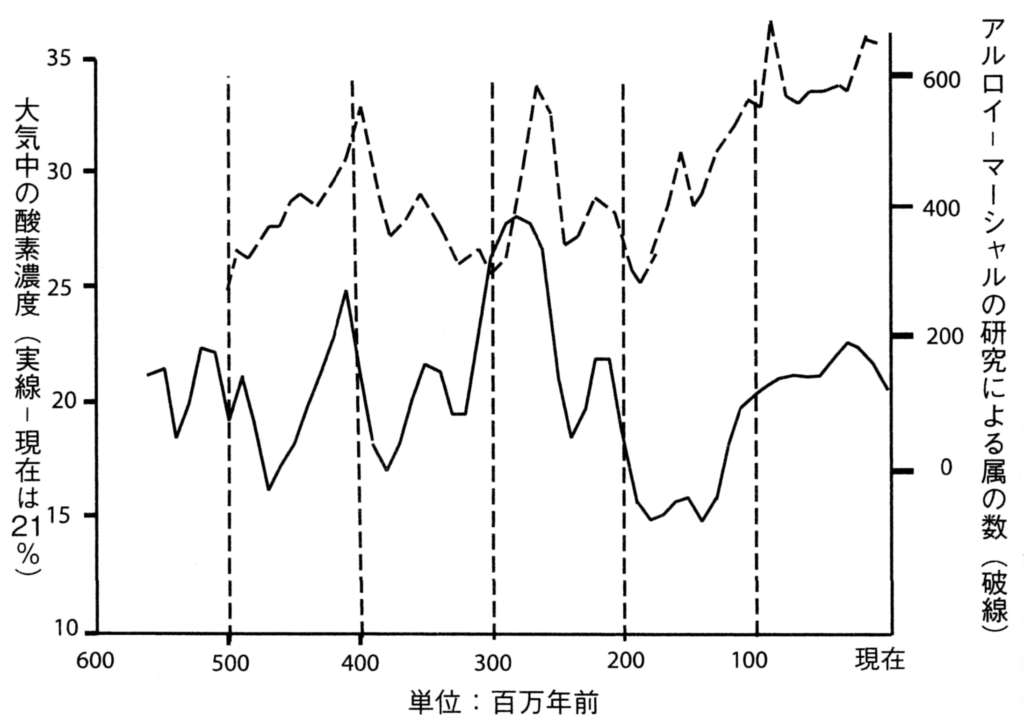
1000万年単位で見た場合、酸素濃度と生物多様性の間には何らかの相関関係が認められるが、特に注目すべき相関性が現れるのは、1000万年ごとの期間内で大気中の酸素濃度の増減を生物多様性の変化と対比した場合である。例えば、2億3000万年前~2億2000万年前の大気中の酸素濃度と、同じ時期の種の多様性の変動を比較すると、明らかな相関関係が存在することが分かる。
つまり、これは偶然ではなく、統計学的に見て非常に高い信頼性を持つ相関であると言える。従って、生物の多様性に最も大きな影響を与える要素は、『酸素濃度』であると考えられる。異なるソースから得られた2つの曲線が相関を示すことにより、科学的な信頼性が向上しているのである。
図表
図107 古杯類のカンブリア紀の化石
恐竜化石グッズの専門店ふぉっしる「古杯類」
(https://www.palaeoshop-fossil.com/catalogue/otherfossil/Archaeocyath004.html)
図108 現代・古生代のサンゴの例
石サンゴ(現代)
フリー百科事典『ウィキペディア(Wikipedia)』「イシサンゴ目」(https://ja.wikipedia.org/wiki/%E3%82%A4%E3%82%B7%E3%82%B5%E3%83%B3%E3%82%B4%E7%9B%AE)
床板サンゴ(オルドビス紀-ペルム紀)
フリー百科事典『ウィキペディア(Wikipedia)』「床板サンゴ類」(https://ja.wikipedia.org/wiki/%E5%BA%8A%E6%9D%BF%E3%82%B5%E3%83%B3%E3%82%B4%E9%A1%9E)
四放サンゴ(デボン紀)
恐竜化石グッズの専門店ふぉっしる「四放サンゴ」
(https://www.palaeoshop-fossil.com/catalogue/otherfossil/Rugosa040.html)
図109 福地層に含まれる層孔虫
ジオランドぎふ「モバイル端末版」
(https://geo-gifu.org/mobile/1_kaseki/kaseki_5_soukouchuu.html)
図110 サンゴのポリプの軟組織と骨格の断面図
環境省、自然環境・生物多様性、サンゴ礁保全の取り組み「参考資料 2- 1 サンゴ礁
の基礎知識」
(https://www.env.go.jp/nature/biodic/coralreefs/reference/mokuji/9932j.pdf)
図111 渦鞭毛藻類
筑波大学 生命環境学群 生物学類 植物分類・系統・進化グループ(https://www.biol.tsukuba.ac.jp/~ken/phycological_images/d/dinoflagellates.html)
図112 ヒラハコケムシ
公益財団法人 水産無脊椎動物研究所・水産無脊椎動物図鑑・コケムシ動物門 (https://www.rimi.or.jp/dobutumenu/c/bryozoa/)
図113 顕生代5回の大量絶滅(ビッグファイブ)
ヘルシスト211号 平成24(2012)年1月10日発行 Special Features 1「生命の「復興力」」
東京大学大学院総合文化研究科教授 磯﨑行雄「「大量絶滅」を乗り越えてきた生命進化」(https://www.yakult.co.jp/healthist/211/img/pdf/p02_07.pdf
図114 顕生代を通しての酸素濃度の変動
ROBERT A. BERNER, JOHN M. VANDENBROOKSAND and PETER D. WARD
“Oxygen and Evolution”,SCIENCE 316 (2007) 557-558
(https://www.science.org/doi/10.1126/science.1140273)
図115 顕生代を通しての無脊椎動物の属数の変遷
John Alroy, et al.“Phanerozoic Trends in the Global Diversity of Marine
Invertebrates”,SCIENCE 321 (2008) 97
(https://www.science.org/doi/10.1126/science.1156963)
図116 酸素濃度と属数の増減の相関関係
「生物はなぜ誕生したのか―生命の起源と進化の最新科学A NEW HISTORY OF LIFE ―THE
RADICAL NEW DISCOVERIES ABOUT THE ORIGINS AND EVOLUTION OF LIFE ON
EARTH」ピーター・ウォード Peter Ward, Ph.D.、ジョゼフ・カーシュヴィンクJoseph
Kirschvink, Ph.D.著、梶山あゆみ訳、株式会社河出書房新社、2016年3月30日 3刷発行
185頁、下のグラフ
