
第四部 生命の進化
第5章 カンブリア爆発と真の極移動【5億4000万 ~ 5億年前】
『カンブリア爆発』に関する最初の主要な証拠は化石から発見された。これらの動物の化石が岩石中に現れるパターンには「四つの波」が確認される。
最初の波が始まったのはおよそ5億7500万年前である。
- これを『アバロン爆発』(注74)と呼ぶ。アバロンとは、カナダ東部のニューファンドランド島の地名で、ここにあるエディアカラ紀の地層から最古の化石群が発見された場所である。
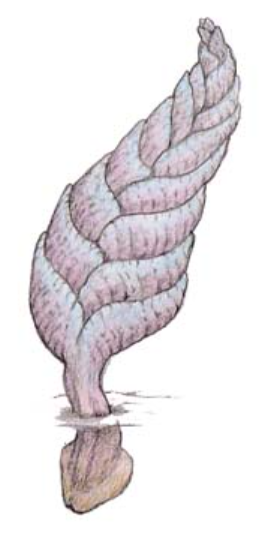
(注74)エディアカラ生物群は、オーストラリア、アデレードの北方にあるエディアカラ丘陵で大量に発見される生物の化石群を指す。これらは約6億~5億5千万年前の先カンブリア時代の生物であり、基本的に目、歯、トゲ、硬い組織などを持たない軟体性であったと考えられている。生存競争の本格化前の「エデンの楽園」を思わせるため、「エディアカラの楽園」とも称される。エディアカラ生物群には数種類の巨大軟体動物が含まれているが、これらは初期動物の進化理解に重要な意義を持つだけでなく、化石の異なる組み合わせは、エディアカラ紀後期の地層の区分と対比のシンボルでもある。エディアカラ生物群の中で最も典型的な化石として海底に固着し生息していた葉状体生物『チャルニア』が挙げられ、その出現のピークは5億7400万年-5億6000万年前のアバロン群集であった。また、最古のエディアカラ生物群であるニューファンドランドの「アバロン生物群」の調査から、エディアカラ生物群の種類はすでにこの時期に全て出現していたとされ、5億7500万年から5億6500万年前にエディアカラ生物群の多様性が形成されたことを『アバロンの爆発』と呼ぶ。しかし、エディアカラ生物群とカンブリア紀以降の生物との類縁関係は不明確であり、両者は対応しないとの見解から、「アバロンの爆発はおそらく失敗に終わった実験だった」と表現されることがある。
第二の波が起きるのは、エディアカラ生物群がほぼ完全に姿を消した時期と重なり、実際の化石ではなく生物の活動の痕跡が正確に保存されている。

- この膨大な数の『生痕化石』は、多細胞生物の活動により形成されたものと考えられている。すなわち『動物』の存在を示すものである。
- 生痕化石の中には5億6000万年前のものも含まれているが、大半は5億5000万年前のものである(注75)。
当時の海底は、活発に動いていた小さな蠕虫のような生命で満たされていたと推定される。
(注75)エディアカラ紀とカンブリア紀の境界は5億4200万年前頃とされている。
第三の波は骨格の出現である。
- 5億5000万年前より少し新しい地層からは、無数の微細な「骨格要素」が見つかっている。これらは非常に小さなトゲやウロコで、炭酸カルシウムでできており、動物の体表をタイルのように覆っていたとされている。
最後の第四の波では、
- 大型の化石動物が出現した。それらは『三葉虫』、二枚貝のような『腕足動物』、トゲを持つ『棘皮動物』、さらには巻貝に似た様々な『軟体動物』などであり、全てが5億3000万年前より新しい地層に含まれている。

ダーウィンの時代には、第一から第三の波についてはほとんど知られておらず、カンブリア紀の始まりは堆積層に最初の三葉虫が確認できる時期とされていた。このように波が連続して訪れた理由は、酸素濃度が過去最高レベルに達していたという意外なほど単純なものであった(<その 8>図87参照)。
四つの波は比較的短期間のうちに化石記録に現れている。最新の年代測定法により、最初の複雑な化石(最古の生痕化石から1000万年前~2000万年後に現われた微細な骨格の化石)が登場する時期は5億4000万年あまり前と特定され、最初の『三葉虫』が出現するのはその2000万年ほど後だったことがわかった。
化石記録に『動物』が登場するのは重要なイベントであり、『カンブリア爆発』と呼ばれている。古生物学者にとって『カンブリア爆発』は、化石に残るほど大型な動物の主要な門のほとんどが最初に誕生したことを意味する。分子遺伝学者にとっては、生命が進化して初めて動物になったことを示している。
その時期については1990年代を通して激しい議論が続いたが、21世紀初頭の高度な分析法を用いた新しい分子研究により、古生物学者が主張していたより新しい時代に動物が現われたことがほぼ確認された。具体的には、
地球上に動物が存在した最も早い時期はおそらく少なくとも6億3500万年前であり、実際には5億5000万年前に近かった可能性があるとされている(注76)。
(注76)『生痕化石』の中には古いもので5億6000万年前のものも含まれているが、ほとんどは5億5000万年前のものである。当時の海底は、活発に動いていた小さな蠕虫のような生命で満たされていたと推定されている(136-7頁参照)。
現在の科学的な理解に基づくと、『カンブリア紀』は約5億4200万年前から4億9500万年前にかけて存在したとされている。ただし、その終了期間が若干早かった可能性もある。なお、
- 動物門(注77)の多数が初めて登場したのは、約5億3000万年前から5億2000万年前という比較的短い期間であった。この事象は生命の歴史全体を通して三番目か四番目に重要な出来事であり、これは専門家の間で共通の認識である(注78)。
(注77)動物門は生物の分類学的階級で、『動物界』の下位に位置する。その例として「節足動物門」、「軟体動物門」、「脊索動物門」などが存在する。
(注78)この現象は『カンブリア爆発』と称され、現在に至るまでの動物の「門」が一気に登場した出来事を指す。
カンブリア爆発よりも重要度が高い出来事として、以下の三つが考えられる:
- 「地球に初めて生命が誕生した」こと
- 「生命が酸素に適応した」こと
そして、
- 「真核細胞が生まれた」こと
しかない。
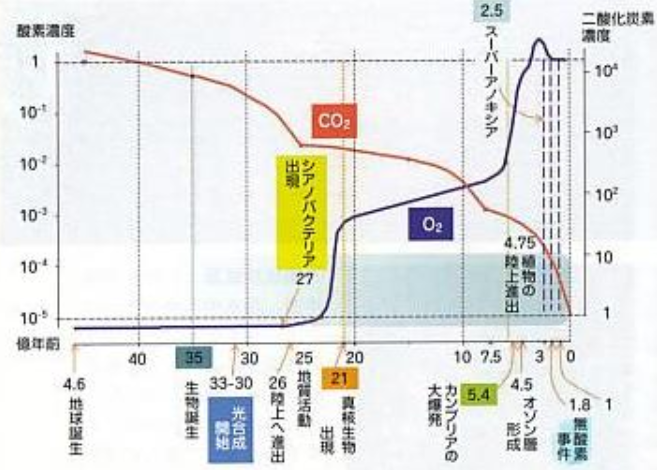
最新の信頼性の高い情報によれば、カンブリア爆発が始まった直後の地球では大気中の酸素濃度は約13パーセントであった(現在は21パーセント)。ところがその後で変動する。この時代、二酸化炭素の濃度は現代の数十倍にも達していた。そこまで高ければ猛烈な温室効果が生じたはずであり、太陽エネルギーが現在より5パーセント低かったとしても十分に温暖化が生じていたと推測される。カンブリア爆発が終わった頃には二酸化炭素濃度は減少したが、動物が初めて誕生した後のどの時代と比べても、地球の気温は高かったと考えられる。高温状態では酸素が海水に溶けにくくなるため、海の酸欠状態はさらに悪化したと思われる。
中国のチェンジャン(澄江)地方で新たに発見された化石埋蔵地には、硬組織と軟組織の両方を含む多種多様な化石が保存されている。この化石群から、動物の門がどのように形成されたのかを推察することができるだけでなく、カナダのバージェス頁岩より前の時代の生命がどのようなものであったのかも推測することが可能である。『チェンジャン』の地層は約5億2000万年前から5億1500万年前に堆積したとされ、一方の『バージェス頁岩』は約5億500万年前以前に形成されたとは考えられていない。これらの地層はおおよそ1000万年の間隔があるため、それらを比較することで動物がどのように多様化していったのかを観察することが可能である。

チェンジャンとバージェスの両地層では、骨格だけでなく軟組織も保存されているため、どの動物がどれほどの数で生息していたのかを比較的正確に把握することが可能である。軟組織が確認できない場合、各動物の種類に応じて存在した数を確実に判断することは困難である。これは、骨格を持たない蠕虫やクラゲのような動物が多数存在していた可能性があるからである。それを考えると、どちらの地域についても動物相の全体像が明らかになっているという事実は驚異的である。現在までに、バージェス頁岩からは5万点を超える化石が発掘されており(チェンジャンではそれより少ない)、デリク・ブリッグズ、ダグラス・アーウィン、フレデリック・コリアーの三人が1994年の著書『バージェス頁岩化石図譜』(大野照文監訳、朝倉書店)で記述した150種の動物のうち、半数近くが節足動物またはそれに類する動物であった。だがそれ以上に興味深いのは個体数である。
化石の数全体では、九割以上が『節足動物』で、次に多いのが『海綿動物』と『腕足動物』であった。これは、より古い時代のチェンジャンと同様に、バージェスの海底でも節足動物が種類でも数でも他の生物を圧倒していたことを示している。

節足動物は無脊椎動物としてきわめて複雑な構造をもっている。にもかかわらず、最も初期の動物化石の中にもすでに数多く存在し、種類も多い。これは、化石記録に初めて姿を現わすより前に長い進化の歴史があったことを物語っている。おそらく体長がせいぜい1ミリ程度の節足動物が海底に多数生息し、泳ぐものや浮遊するものも多く存在したと推測される。
バージェス頁岩の動物相や植物相についての多くの文献は、体が柔らかく美しい生物の紹介に大部分のページを割いている。しかし、驚くべきことに、
バージェス地層で最も一般的な化石はこれらの風変わりな分類群のものではなく、『三葉虫』であったという事実が明らかになっている。
- 三葉虫と、その数は少ないものの多様性を持つ「節足動物」が、バージェス化石群の大部分を占めているという事実は注目すべきである。個体数においても種の数においても、また体制の種類の数においても、この事実が当てはまる。
- 従って、「節足動物」がカンブリア紀の動物の中で最も繁栄した群と考えるのが適切である。
- では、その成功には、節足動物の主要な体制である『体節制』がどれくらい貢献したのだろうか。
地球上の動物の中で最も多様化が進んでいるのは「体節」をもつものであり、その大部分は「節足動物」である。
昆虫をはじめとする多種多様な節足動物は、多数の「体節」を有し、その繰り返し構造を形成している。個々の体節はそれぞれ固有の機能を果たす。
- 節足動物に共通する特徴は、全体が外骨格に覆われていることである。この外骨格は消化管の中にまで侵入している。なお、外骨格は成長しないため、定期的に脱皮を行い、新たなものと交換しなければならない。
- 分化の進んだ頭部、胴体、後端部がどういう比率になっているかは、種によって異なる。
一般に付属肢は特殊化している。陸生節足動物の場合は、個々の付属肢は通常一本であり、それは非常に大きいものであるが、海生のものでは付属肢が二つの枝で構成されている。内側の枝は歩脚であり、外側の枝は鰓である。これらを「二枝型付属肢」(図98、注79)と呼ぶ。
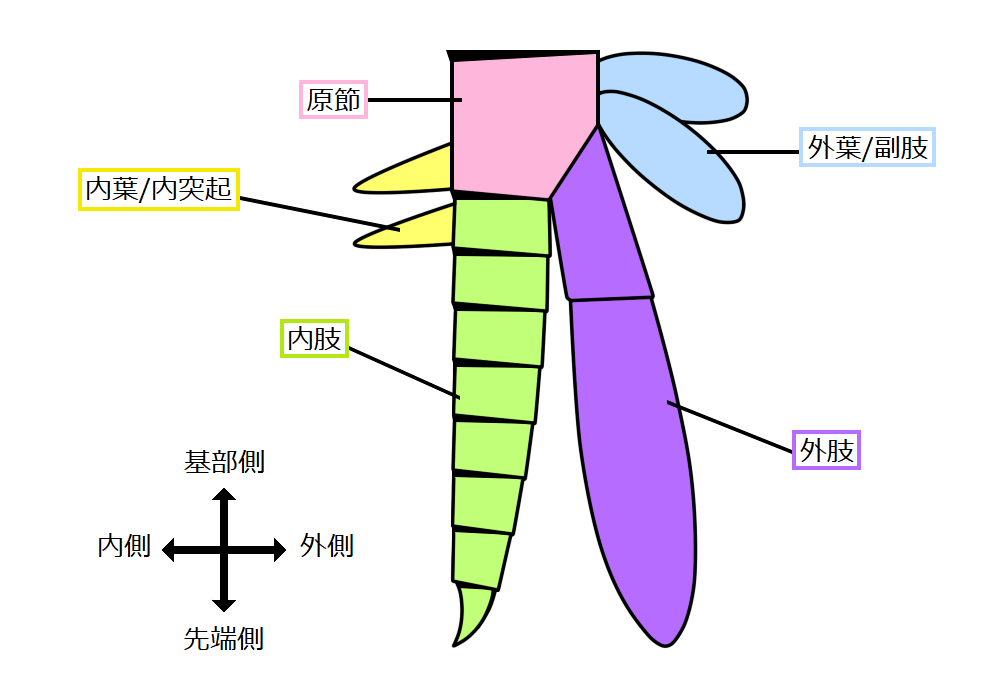
(注79)六脚類、鋏角類や多足類のほとんどの節足動物は、関節肢に分岐した肢節はなく、これは「単枝型付属肢(単肢型付属肢)」と呼んでいる。一方、多くの甲殻類の関節肢は、第1触角以外では基本として途中から2つに枝分かれ、「二叉型付属肢(二肢型付属肢)」と呼ばれる。この場合、外側の分岐は「外肢(枝肢)」、内側の分岐は「内肢(主肢)」、分岐より前の残り全ての肢節は「原節」として区別される。しかし甲殻類の中でも、内肢と外肢のうちどちらの一方(主に内肢)だけが発達して、外見上は単枝型に見える例が多く、十脚類(カニ・エビなど)や等脚類(ダンゴムシ・ワラジムシなど)の胸脚がその代表例である。
外骨格は柔らかい部分を防具のように保護し、その保護が主な機能であると考えられている。しかし、この特性により重大な影響が生じる。それは、体のどの部分からも酸素を受動的拡散によって取り込むことができないのである。酸素を摂取するために、初期の節足動物(全て海生)は「特殊化した呼吸構造」や「鰓」を発達させる必要があった。
なお、体節を持つ特性は節足動物だけが有するものではない。『環形動物』(注80)にはすべて体節があるし、軟体動物の『単板綱』(注81)などにもある程度の体節が確認できる。
(注80)多くが原則として体節制をもち、体は環状の柔らかい体節に分かれている蠕虫状の動物。ミミズ、ヒル、ゴカイなど。
(注81)軟体動物門単板綱の貝の総称。貝殻亜門 (腹足類,頭足類,二枚貝類と掘足類を含むグループ) の祖先と考えられる類で,ネオピリナ(単板類の現生種で,「生きている化石」といわれる)に代表される。
体節は動物の進化の早い段階で現れ、それが初期の動物化石、特にカンブリア紀の三葉虫の化石の一般的な特徴である。
生物学者のジェームズ・ヴァレンタインは2004年の著書『門の起源について』の中で、進化をめぐる大きな謎の一つについて考察している。その謎とは、
- カンブリア紀になぜ節足動物の数と種類がこれほどまでに多かったのか
である。一口に節足動物といっても体のつくりは多種多様である。系統の異なる複数の生物群が、収斂進化を通して似通った姿になったにすぎないからである。だが一つ共通点がある。それは、体節ごとに二枝型の付属肢をもち、それぞれが歩脚と長い鰓を備えているという事実である。
なぜ底生生物(水域に生息する生物のうち、主に、水底を構成する土砂や岩などを生息の基盤として利用する生物の総称)は体節制を選んだのかと問いたくなるが、そもそも体節に分かれていると捉えること自体が間違っているのかもしれない。ヴァレンタインらによれば、少なくとも環形動物は体節ごとに体腔もおおむね区切られているのに対し、節足動物の場合はそこまでではない。むしろ体節が「反復して現れる」と表現するのが適切である。
この独特の体制が生まれたのは、移動の必要性に応えるためであったとヴァレンタインは推測している。「節足動物の体節制が、体を動かすメカニズム、とくに移動と関連しているのは確かであり、それを神経と血液が支えている」。この種の体制が、移動を容易にするためのものであることは疑いようがない。(注82)
(注82)中枢からの信号が分節化した神経系を連続的に刺激し少しづつ位相のずれた継続波を各分節に起こさせることで、神経系に依存した移動を促進、加速することができる。これは波状の遊泳、蠕動運動、潜穴行動、匍匐運動、付属肢による移動に効果的である
しかし、体の構造がこのようになった結果、体節ごとに鰓が繰り返される構造になった。個々の鰓は小さく、体節の下で最適な向きに調節できる。この配置により、羽根状の鰓に能動的に水を通すことで、鰓に接触する酸素分子の数を増加させることが可能である。
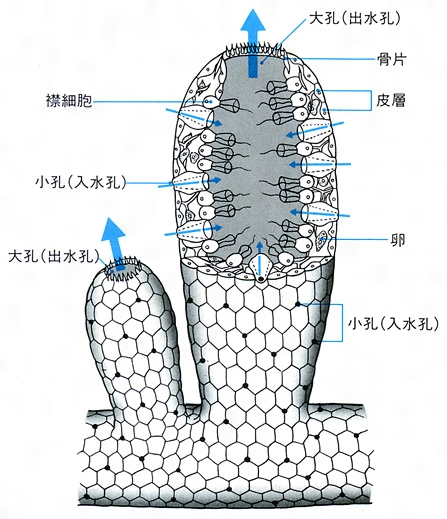
カンブリア紀の最も古い堆積物からたくさん見つかる動物はもう一種類ある。『海綿動物』である。「刺胞動物」(クラゲ・サンゴ・イソギンチャクなど)と同様に、海綿動物には呼吸器官がない。内部に空洞が存在し、その周囲には多数の袋状の構造が配置されている。刺胞動物に似ているが、整然とした構造を持たず、実質的な体組織をもっていない。どの袋も、その体積に対して表面積が非常に大きい。無数の単細胞生物が集合したような構造で、その全体が海水と接触していると言える。しかし、これだけの特性があるとはいえ、海綿動物の酸素取り込み方式は極めて効率的である。食物摂取を主に担当する「襟細胞」が大量の海水を体内に引き込む。一部の専門家によると、海綿動物が一日に取り込む海水の量は自分の体積の一万倍にも上るという。このため、海綿動物は極めて低酸素の環境下でも生存することが可能である。酸素濃度が極めて低い水中でも、その分を補うための大量の水を体内に引き込むことが可能だからである。
カンブリア爆発を引き起こした物理的・化学的原因
古くからある生物学の柱の一つで、近年再評価され、新しい学問といっていいような重要な成果を生み出している分野がある。『進化発生学』、通称『エボデボ』(「発生進化生物学Evolutionary Developmental Biology」)である。この分野は過去10年で目覚ましく発展し、カンブリア爆発に関する多くの研究が発表されてきた。
エボデボ界を代表する研究者の一人ショーン・キャロルは、2005年の著書『シマウマの縞 蝶の模様―エボデボ革命が解き明かす生物デザインの起源』(渡辺政隆/経塚淳子訳、光文社)の中で、以前は扱いが難しいとされていた進化生物学の問題が現在ではより理解可能になっていることを述べている。
その問題とは、「新奇性」がどのように発生するかという点である。進化の過程で、それまで存在しなかった形質が比較的短期間で出現することがあり、この現象は従来のダーウィンの進化論では説明が難しかった。翼や脚の出現、節足動物の体節の形成、またはカンブリア爆発の特徴である体の大型化といった急激な進化がそれに当たる。
キャロルは著書の中で、進化における劇的な変化を新たな切り口からうまく説き明かす四つのポイントを挙げている。
キャロルが「革新のための秘訣」と呼ぶものの一つ目は、
- 既存のものを利用する
である。新しいものを生み出すためには、新たな装置や道具を作る必要はなく、既存のものを利用することが最も効率的である。
二つ目と三つ目はダーウィンも認識していた概念で、
- 多機能性
- 反復性
である。多機能性とは、既存の形態や構造、あるいは生理機能を利用して、その本来の機能に加えて新たな機能を付与することを指す。反復性とは、何らかの構造が複数の部分で構成されており、それらが共同して全体の機能を果たす場合を指す。一部の構造に新しい機能を付与し、残りの部分が従来通りに機能するなら、全く新しい構造を一から作り出すよりも容易に新たな進化が可能となる。
その典型例が『頭足類』(「軟体動物門頭足綱」に属する動物の総称で、イカ、タコ、オウムガイ、コウモリダコや絶滅したアンモナイト等が含まれる)の泳ぎと呼吸である。頭足類は大量の水を吸い込んで鰓に通しており、多くの無脊椎動物と同様、水の吸入と排出は別々の「管」を用いて行う。これは酸素を多く含む水を無駄に排出しないようにするためである。ただし、この管の構造をわずかに変化させるだけで、新たな移動手段が得られる。呼吸と移動のエネルギー消費は以前と変わらないまま、同じ量の水を利用して呼吸と移動の両方を実現できるようになったのである(注83)。

(注83)イカの場合、呼吸のため外套膜の中に取り込んだ海水を、勢いよく吐き出すことにより推進力を得る重要な推進器官が「漏斗」である。漏斗を前後左右に曲げることによって、前進、後進、自由自在に進むことができ、また、外敵の目を眩ますための墨もここから排出される。
四つ目の秘訣は
- モジュール性
である。節足動物はもちろん、人間を含む脊椎動物も、体が体節に分かれているという意味ではすでに「モジュール性」をもっていると言える。節足動物の体節からは付属肢が伸びており、それぞれが途方もないほどに改造されて、摂食、交尾、移動など様々な機能に適応する構造を持っている。
節足動物はまさにアーミーナイフのようで、特定の機能に特化した付属肢を個々の体節が備えている。これは人間を含む脊椎動物にも当てはまる。元々は原始的だった手足の指は、陸を歩く、水中を泳ぐ、空を飛ぶなど多様な動きが可能になるまで進化してきた。
実際、このような構造は、柔らかいパテのように形を変えられることが明らかになっている。これはその根底にある『遺伝子スイッチ』のシステムが存在するためである。このスイッチが位置しているのは発達中の胚であり、これが後に節足動物や脊椎動物の様々な付属肢になる部分と同じ場所である。スイッチは重要な役割を果たしており、いつどこで成長するべきかを体の各部位に指示している。
節足動物の体が頭部、胸部、腹部という順番になっているのは、それぞれがまずは染色体上で、次いで胚自体の中で、既に完全に同じ順番で並んでいたからである。これは非常に驚くべき発見であり、この仕組みの大部分を担当しているのが、エボデボの分野で王者とも呼べる『ホメオティック遺伝子』(注84)である。この遺伝子を持つのは節足動物だけではなく、名前は違うが同等の遺伝子が他の分類群の生物にも存在している。
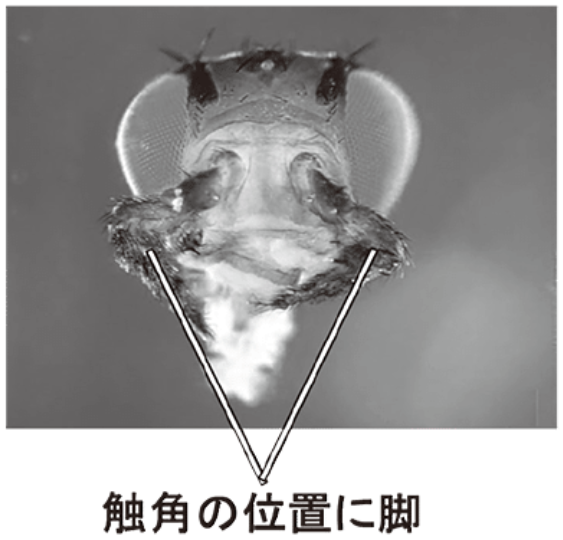
(注84)ホメオティック遺伝子とは、生物が正しく発生、分化するためのキーとなる遺伝子で、動物の胚発生の初期において組織の前後軸及び体節制を決定する遺伝子である。この遺伝子は、胚段階で体節に関連する構造(例えば脚、触角、目など)の適切な数と配置について決定的な役割を持つ。例えば、ショウジョウバエの体節の特性を制御する遺伝子に変異が起きると、そのショウジョウバエでは触角の代わりに脚が生えるという奇妙な現象が起こる。体節の性質が別の体節の性質に変化する現象を「ホメオーシス」というが、このように体節の特性を制御する遺伝子が「ホメオティック遺伝子」である。
前述の通り、カンブリア紀の始まりは現在、約5億4200万年前と特定されている。その起点となるのは、地層中において動物の移動の痕跡が初めて確認される場所である。具体的には、特定の生痕化石により、動物が泥を縦に掘った跡が発見される地点である。しかしその後の1500万年間で新しい体制が誕生した形跡はほとんど見られない。少なくとも化石記録にその証拠が残されていないのである。動物の爆発的な多様化を証明する最初の手がかりは、最近になってチェンジャン(澄江)から提供された。前述した通り、この地域の約5億2500万年前~5億2000万年前の地層からは驚くべき化石群が見つかっている。時代は古いものの、柔らかい部分も保存されている点でバージェス頁岩に似ている。
「エボデボ」の分野から生まれた多くの新発見は、『カンブリア爆発』の謎を解く上でも利用されてきた。その中でも最も重要な謎は、現存する多種多様な動物の『門』と『体制』がいつ、どのように始まったのかという問いである。

チェンジャンでもバージェス頁岩でも、動物群を数で圧倒しているのが『節足動物』(図102)である。その種類は実に多岐にわたる。すぐに節足動物は地球上で最も多様な動物群になり、その地位を保持し続けることになる。現在では、甲虫だけでも約3000万種が存在すると推定されている。
エボデボの分野はその理由を明らかにしている。多様な体制の中で、節足動物のものほど容易かつ短期間に、しかも劇的に変化できるものはない。キャロルが提唱した特徴を完全に持っているからだ。すなわち、
- モジュラー構造の部分から成り立ち、
- 形態の繰り返しがあるので新しい機能を担わせやすく、
- 体節の特定領域を速やかに変容させる一連のホメオティック遺伝子を保有している。
かつては、新種の動物が誕生するためには新しい遺伝子が必要だと考えられていた。原始的な海綿動物やクラゲは、より複雑な節足動物に比べれば遺伝子の数が少ないはずだから、全節足動物の共通祖先は新しい遺伝子、すなわち新しいホメオティック遺伝子を何らかの方法で追加したに違いないと考えられていた。何故なら、いつ、どのように発達するかを体の各部位に指示するのがホメオティック遺伝子の「スイッチ」なのだからだ。しかし、それは誤りであった。
キャロルらが解明したのは、節足動物の最後の共通祖先が新しい遺伝子を進化させたわけではないという事実だ。
- 遺伝子はすでに存在していた。
- 節足動物がのちに驚くべき多様化を遂げるのは、既存の遺伝子を利用してのことだったのである。
「形態が進化するうえで問題になるのは、どんな遺伝子を持っているかではなく、それをどう利用するかである」とキャロルは指摘している。
10種類のホメオティック遺伝子さえあれば、節足動物を一新し、多様化させることが可能である。そのメカニズムが明らかになったのは、各ホメオティック遺伝子から生成される特有のタンパク質を比較し、それらが初期胚のどの部位に存在するかを調査した研究からである。かつて、節足動物のある遺伝子が脚を生成する指令を持つと考えられていたが、それは誤りであった。
ホメオティック遺伝子の主な役割はタンパク質の合成である。そしてそのタンパク質が、初期胚の特定の領域の成長を促進または停止する手段となる。これらのタンパク質の中には、特化した付属肢の生成に関わるものも存在する。もし胚の別の位置にこのタンパク質を移動させたとすると、そこから生じる産物も移動する。結果として、本来なら体の特定の部位から生えるはずの脚が、全く新しい場所から生じる可能性がある。ただし、その場合、実際に脚が形成されるよりも前にそのタンパク質が移動している必要がある。
- 何れにせよ、ホメオティック遺伝子が生成するタンパク質の位置を胚の中で変えることにより、新たな形態の生成が可能となる。
- 節足動物の胚内でこのプロセスが発生した結果、我々が現在観察する多種多様な種類の進化が遂げられたのである。
節足動物の形態には数千種類、あるいは数百万種類も存在すると推定される。それらは全て、10個の遺伝子を用いて生み出されている。体節の反復性が節足動物の特徴であり、各体節が適切に特化するためには、それぞれが異なるホメオティック遺伝子からの指示を受ける必要がある。
カンブリア爆発と真の極移動
生命の歴史の中で『カンブリア爆発』は特筆すべき重要なイベントである。しかし、近年まで理解が進まなかったのは、カンブリア紀の年代が正確に特定されていなかったからである。各種の発見や仮説、議論を経て、
カンブリア紀の開始は三葉虫の出現という目安から、より古いイベント、すなわち縦に掘られた生痕化石を基準とすることとなった。結果として、カンブリア紀の始まりは約5億4200万年前と判明した(<その2 図17>参照)。
カンブリア紀は他に類を見ない時代であり、驚異的な進化と新たな展開が見られた。しかしそれだけでなく、その後の研究で他にも独特な特徴が明らかになっている。原生代(エディアカラ紀)とカンブリア紀の地層の境界で炭素の同位体を調査した結果、非常に奇妙な現象が確認されたのである。
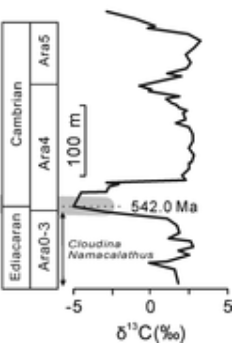
- 数十万年にわたり大規模な変動が見られ、これを『カンブリア紀の炭素循環変動』と呼ぶ。その変動幅は大きく、地球上の全バイオマスを200万~300万年おきに全焼するのに匹敵する。何らかの原因で軽量の炭素がメタンとして大気中に大量に排出され、それに伴い様々な温室効果が顕著となった。地球が短期間に過熱する周期が存在した可能性がある。
さらに奇妙な特徴として、カンブリア紀には非常に大規模なプレート運動が発生したことが知られている。この運動は古地磁気を調査することで追跡可能であり、特定の岩石がかつて存在していた緯度やプレート運動の方向も割り出せる。当時、古地磁気に関する多数の新しい分析から、一見すると信じ難い結果が得られていた。
全ての大陸が地球表面を高速で移動したか、あるいは自転軸はそのままに地球全体が急速にずれたかのどちらかである。北極と南極は従来の位置にありながら、その下にある地球自体が動いたのである。
この結論は驚異的であった。
同様のプロセスは火星や月、さらには多くの衛星や小惑星で何十億年も前から起きており、これらの天体は「自転軸の位置を大きく変更する」ことがある。地球でも同じ現象が起きたら、生命に計り知れない影響が及んだ可能性がある。この可能性についての調査はまだ始まったばかりだが、生命の歴史に対する私たちの理解を大きく変える可能性がある。
- かつて月および火星において、元々は赤道でない位置に新たな質量が加えられ、それが結果的に赤道の位置に落ち着いたという事態が起きたことが確認されている。
具体的には、火星に広がるタルシス地域は、膨大な量の重い溶岩で構成されている。この溶岩は、形成された天体に後から追加された鉛片のような存在であり、回転するフリスビーやボールに見立てることができる。さらに詳しく説明すると、タルシス地域は太陽系内で最大の「重力異常」(注85)を示し、火星の赤道上に位置している。月においてもアポロ計画以前の調査から、平原に玄武岩による質量の集中が赤道上に存在することが確認されていた。火星と月の両天体にはプレートテクトニクスが存在しないため、この現象の理解は容易である。
(注85)重力の実測値から標準重力値への偏差を指す。「標準重力」とは、自転する地球楕円体上での重力を指し、赤道上で最小、両極上で最大となるような緯度の関数で表現される。現在広く使用されている標準重力の式は、1979年に国際測地学地球物理学連合(IUGG)の総会で採用が決定され、これを「正規重力式」と呼ぶ。実際の重力測定は高度が異なる点で行われるため、重力異常に対するいくつかの補正が必要である。
この自転軸の位置変化のプロセスは『真の極移動』(注86)と呼ばれる。
(注86)『真の極移動』については<物語り その8>の注73、図91で説明したが、再度以下に整理して、記載する。
- まず、『極』には『地理極』および『磁極』の二つが存在する。『地理極』は、自転する天体(惑星、小惑星、衛星など)における自転軸(地軸)と地表面との2つの交点、つまり「北極点」と「南極点」を指す。北極点は赤道から北に90度、南極点は南に90度の位置にある。
- 全ての天体は地理極を持っており、地球のように天体が磁場を発生している場合は、地理極とは別に『磁極』(「北磁極」と「南磁極」)も存在する。
- 古地磁気学により、火成岩生成時の偏角と伏角を取得することが可能だが、偏角は極の方向を示し、伏角は緯度で決まる。つまり極からの距離で決まるため、当時の極の位置を知ることができる。ここで得られるのは<厳密には極ではな「磁極の位置」であるが、数千年以上の時間スケールで取得された複数のサンプルを平均化すれば、「磁極の分布の中心は極とほぼ一致する」。
- 『極移動』とは、地質学的時間スケールで固体地球またはその一部に対する磁極(磁北極・磁南極)の移動を指す。
- なお、極が実際には動いていなくても、大陸が移動すると、大陸から見た極の位置は変わる。大陸移動説以前には、これを実際に極が移動したと解釈されており、その名残で『見かけの極移動 (apparent polar wandering = APW)』と呼ばれる。一方、大陸移動を補正すると、地殻全体に対する極の移動が残る。これを『真の極移動 (true polar wandering = TPW)』と呼ぶ。大陸移動、氷床の盛衰、大規模な火山活動、大規模な天体衝突、地球内部の質量分布の再編などにより、固体地球の質量分布が変化し、慣性能率テンソルが変化すると、「真の極移動」が起こる。
地質学的に考えると、惑星上の質量が急激に変化するような事象は様々に考えられる。例えば、大型の小惑星や彗星の衝突や、地球内部のマグマの地表噴出がある。同様に、プレート運動に関連する構造(「拡大中心」や「沈み込み帯」、<物語り その2>図21参照)が現れたり消えたりすることでも、大幅な質量の変動が起こりうる。新たに増加した質量が何らかの方法で浮かんで漂うのではなく、能動的に維持されている限り、地球上で真の極移動を引き起こすには十分である。その構造が消失する場合も影響が生じる。沈み込み帯や拡大中心が消滅するとすれば、一つの大陸が移動して別の大陸と衝突する時である。衝突する二つの大陸間の海底に拡大中心や沈み込み帯が存在していれば、衝突によって破壊される。その結果、地表の質量が減少し、自転軸の位置が変化することになる。
大陸の急速な移動が何らかの原因で進化のペースを加速させたと考えることは理解できる。そのメカニズムはいくつか発見されており、カンブリア紀について知られている情報の断片をうまく結びつけて説明してくれる。
- 一つには、大陸が高緯度に位置していると、海底や永久凍土の中に凍ったメタンを蓄積しやすい。これを「メタンハイドレート」という。その地域が赤道に向かって移動すると、気温が徐々に上昇し、その結果として温室効果ガスが大気中に放出される可能性がある。これが環境の温暖化につながり、温暖化が生物の代謝を促進すると、進化や種の多様化が速く進む傾向にある。
ジョゼフ・カーシュヴィンクらが科学雑誌における発表でこのメカニズムを明らかにした際、彼らは論文のタイトルとして『カンブリア爆発を引き起こしたメタンの導火線』を採用し、種の急増の大きな要因の一つが熱循環であった可能性を提唱した。また、彼らは炭素同位体の異常な移動もこの現象が一因となった可能性を示唆した。
- 赤道地方における生物多様性の自然発生も確認されている。著者らの研究パートナーであるイェール大学のロス・ミッチェルが真の極移動中の大陸の動きを調査したところ、新たに登場した動物群のほぼすべてが赤道へ向かう大陸の先端部分で生まれたことが明らかになった。逆に、高緯度地方へ移動した地域では新たに誕生した生物がほとんどなかった、あるいは非常に少数であった。
緯度が関与しているという理論は極めて単純なものであるが、生物多様性の増加を適切に説明している。自然がホメオティック遺伝子を介して多種多様な体制を試行している際にそのような現象が起こると、多様性の増進が促進されるであろう。
なお、「真の極移動」はカンブリア爆発にのみ当てはまるのではなく、大量絶滅が起きるメカニズムの説明にもなる。そうした絶滅の一つがカンブリア紀とカンブリア爆発を終わらせ、バージェス頁岩の奇妙奇天烈動物をほぼすべて消し去ってしまった。
カンブリア紀の終焉 - 顕生代初の大量絶滅
先カンブリア時代末期に生息していた生物は、動かなかったり、漂っていたり、あるいは比較的大型であっても構造が単純な動物であった。しかしながら、カンブリア紀の終わりには世界中の海には数も種類も豊富な動物が存在していた。だが、なぜカンブリア紀に終わりが訪れたのかという問いについては、従来の理論を覆す研究結果が得られている。
『大量絶滅』とは、短期間で個体と種の両方が大量に死滅することであり、その規模は事象により異なる。特に大規模なものとしては『ビッグファイブ』と呼ばれる五度の大量絶滅がある。それぞれの場合で種の50パーセント以上が失われた。しかし、それほど壊滅的ではないものの、絶滅は他にも複数回発生している。その中でも特に有名なものは、カンブリア紀を終わらせた大量絶滅(注87)である。
(注87)古生代のカンブリア紀末(約4億8800万年前)に大量絶滅が発生し、三葉虫、腕足類、コノドント(注88)が激減した。また、カンブリア紀には約5億1700万年前および約5億200万年前にも大量絶滅がたて続けに起きている。
(注88)カンブリア紀から三畳紀(6億年前から1億8千万年前)の地層から発見される歯状の微化石である。一般に大きさは0.2ミリ~1ミリ程度。動物体の一部の化石で、その正体は長く謎のままだったが、世界中で発見され示準化石として様々な成果を挙げた。発見された化石が魚の歯に似ていたため、ラテン語で「円錐状の歯」を意味する「コノドント」と命名された。現在ではクリダグナサスなどの原始的脊椎動物の歯であると考えられている。
カンブリア紀末期の絶滅は、一度だけでなく比較的小規模な絶滅が3~4回発生しており、主に三葉虫や腕足類などの海洋無脊椎動物が影響を受けた。三葉虫の中でも初期に現れたオレネルスなどは、この時期に完全に絶滅した。そして、三葉虫全体の特性が変化した。カンブリア紀の三葉虫は多数の体節に分かれ、目は原始的で、防御用のトゲなどの構造が体に付属していなかった。さらに、現代のダンゴムシが危険を感じたときのように、体を丸めて硬い玉にする能力も持っていなかった。しかし、続くオルドビス紀に入ると新たな進化の波が訪れ、三葉虫は体制そのものを変化させた。ほぼすべての種が体節の数を減らし、外骨格を厚くして割れにくくなり、目の機能を向上させ、防御構造を発達させ、そして何よりもダンゴムシのように体を丸くする能力を得た(注89)。
(注89)『三葉虫』という名前の由来は、胴体部分が中央の軸、その左右の房状の部分と三つに分かれ、体が「三つの(tri)」、「葉または房(lobe)」 からなる「石(ite)」であったため、『三葉虫(trilobite)』という名前がつけられた。身体を覆うこれらの硬い殻に守られ、その下には何本もの脚がダンゴムシのように生えていた。

三葉虫類は、カンブリア紀(約5億4200万年前)からペルム紀末(約2億5100万年前)まで、世界中の海で広く繁栄し、その種類は実に約1,500属10,000種くらいにもなるのではないかと考えられている。大きなものは70cmほどにもなり、逆に、小さいものでは成体でも5mm程度という種も見つかっている。目玉がかたつむりのように飛び出した「アサフス」に見られるように、三葉虫は最初に「視力」をもった動物のひとつと考えられている。多くの種ではトンボの目のような複眼がみられるので、ものの形が認識できるほどに視力が発達していたと考えられる。そして、もう一つの特徴は「硬い殻」である。三葉虫が出現したカンブリア紀以前にいた生物として「エディアカラ生物群」が知られているが、それらの大部分は硬い殻など持たず、やわらかいフニャフニャの姿で生きていたと考えられている。
なぜ、三葉虫は、視力、硬い殻が発達したのだろうか。それは、この時代に三葉虫を餌とする捕食者が現れたことと深い関係がありそうである。視力をもって敵をすばやく察知して逃げ、硬い殻で身を守ることで、三葉虫は古生代末期まで繁栄することができたのだと考えられる。三葉虫の硬い殻や、背中のトゲなどは、強力な敵(アノマロカリスやウミサソリなど)からの攻撃を防ぐのに有利だったと考えられる。三葉虫の背側の殻は「カルサイト」(「炭酸カルシウム」が結晶化した鉱物『方解石』)でできていたと考えられ、よく化石として残るのだが、中には、殻の一部が他の動物に食べられた跡のように楕円状に欠けている化石や、ダンゴムシのように丸まって身を守っているかのような状態の化石が発見されている。これらは、カンブリア紀の海底で壮絶な食うか食われるかのバトルが始まっていたことを想像させる。(東北大学総合学術博物館・展示案内・ミニ標本案内http://www.museum.tohoku.ac.jp/exhibition_info/column/trilobite.html)より)
カンブリア紀末の絶滅に関しては、従来、高温と低酸素、そして動物相(ある特定の地域と時間における動物を表す集合的な用語)の変化によるものと考えられていたのに対し、その正反対を示唆する新たなデータが次々に見つかってきた。
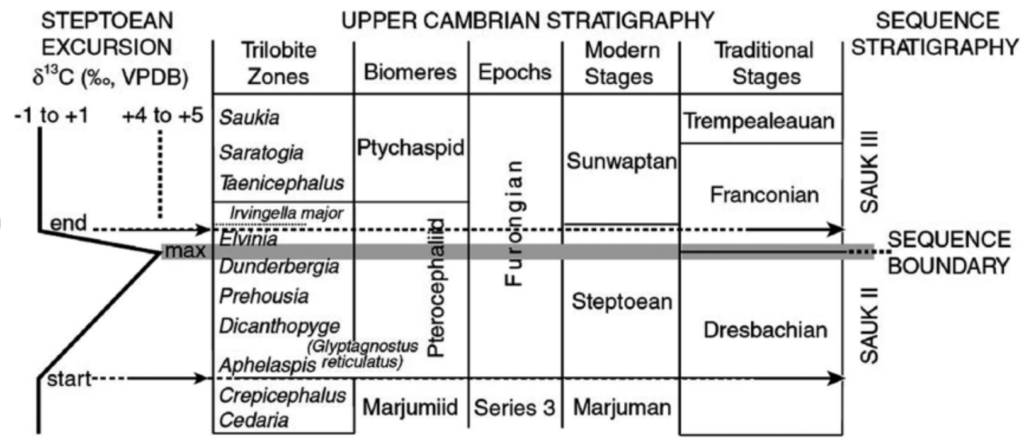
水が温かいどころか冷たかったことと、海底への有機物の大規模な埋没が起きていたことを示す証拠である。このプロセスにより酸素濃度は急上昇したのである。現在、この変化は『SPICE(「後期カンブリア紀の正の炭素同位体変動」の略)』(注90)と呼ばれている。
(注90)The Steptoean(フロンジアン世(497~485.4百万年前)の地質年代層序区分前期の『期』) Positive Carbon Isotope Excursion (SPICE) was a geological(地質学の) event which occurred about 494.6 million years ago near the beginning of the International Furongian Epoch(世) of the Cambrian Period(紀). The SPICE event was a positive shift in carbon isotope (δ13C)(注91)values which lasted for around 2 to 4 million years. This shift is interpreted to be a global disturbance in the carbon cycle, affecting both the ocean and atmosphere. Regional sea level changes, a rise in sea water temperatures, and trilobite(三葉虫) and brachiopod(腕足動物) extinctions are associated with the SPICE event, although the exact mechanism(s) driving these events is still unconfirmed.(https://en.wikipedia.org/wiki/Steptoean_positive_carbon_isotope_excursion)
(注91)『炭素同位体比』については、<物語り その7>(注57)を参照のこと。
他の時期の大量絶滅は、通常、酸素濃度の低下に伴って発生している。しかし、SPICEの場合は、短期的な酸素濃度の上昇と連動して発生したという特徴がある。これは、火山の噴火と同時期の急速な大陸移動、つまり真の極移動が引き金であった可能性が指摘されている。数百万年をかけて多くの陸地が熱帯へ移動した結果、炭素の埋没量が増え、大気中の酸素濃度がこれまでにないレベルに上昇したのである。
(注92)上記のとおり、火山の噴火と同時期に真の極移動を引き起こし、数百万年の間に多くの陸地が熱帯へ移動した。その結果、炭素の埋没量が増え、大気中の酸素濃度が過去最高レベルに上昇するとともに、海中の酸素濃度を低下させた。これが原因となり、海中の大量絶滅と「正の炭素同位体変動(δ13C)SPICE」を起こしたのである。
これらの地球環境の変動が、カンブリア爆発の後の大規模な「適応放散」(注93)への準備を整える契機となったかもしれない。多量の酸素を必要とする生態系、すなわちサンゴ礁がその一つである。サンゴ礁はSPICE直後に登場し、次の地質時代である『オルドビス紀』への道を開いたのである。
(注93)同一起源の生物群が、さまざまな異なる環境に最も適応する形態的または生理的な特性を発展させて、多数の系統に分化すること。
図表
図92 チャルニアの想像図
古世界の住人
(https://paleontology.sakura.ne.jp/tyarunia.html)
図93 円筒形巣穴の生痕化石
フリー百科事典『ウィキペディア(Wikipedia)』「生痕化石」(https://ja.wikipedia.org/wiki/%E7%94%9F%E7%97%95%E5%8C%96%E7%9F%B3)
図94 三葉虫・腕足動物・棘皮動物・軟体動物の化石例
フリー百科事典『ウィキペディア(Wikipedia)』「三葉虫」
フリー百科事典『ウィキペディア(Wikipedia)』「腕足動物」
化石販売の化石セブン
ナショナル ジオグラフィック・ニュース・大発見!カンブリア紀の新たな化石群、中国(https://ja.wikipedia.org/wiki/%E4%B8%89%E8%91%89%E8%99%AB, https://ja.wikipedia.org/wiki/%E8%85%95%E8%B6%B3%E5%8B%95%E7%89%A9,
図95 大気中の酸素・二酸化炭素濃度の変遷
三村芳和著「酸素のはなしー生物を育んだ気体」、中公新書(2007年12月)
図96 バージェス頁岩の節足動物例 アノマロカリス・カナデンシス
フリー百科事典『ウィキペディア(Wikipedia)』「アノマロカリス」(https://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%8E%E3%83%9E%E3%83%AD%E3%82%AB%E3%83%AA%E3%82%B9)
図97 節足・海綿・腕足動物の例
(a)節足動物・オパビニア
フリー百科事典『ウィキペディア(Wikipedia)』「オパビニア」(https://ja.wikipedia.org/wiki/%E3%82%AA%E3%83%91%E3%83%93%E3%83%8B%E3%82%A2)
(b) 海綿動物@沖縄サンゴ礁
岐阜大学教育学部理科教育講座地学教室
(http://chigaku.ed.gifu-u.ac.jp/chigakuhp/html/kyo/seibutsu/Doubutsu/sango/index.html)
(c) 腕足動物・オルチス目
フリー百科事典『ウィキペディア(Wikipedia)』「腕足動物」(https://ja.wikipedia.org/wiki/%E8%85%95%E8%B6%B3%E5%8B%95%E7%89%A9)
図98 二枝型付属肢の模式図
フリー百科事典『ウィキペディア(Wikipedia)』「関節肢」(https://ja.wikipedia.org/wiki/%E9%96%A2%E7%AF%80%E8%82%A2)
図99 海綿動物の模式図
コトバンク、日本大百科全書(ニッポニカ)「海綿」(https://kotobank.jp/word/%E6%B5%B7%E7%B6%BF-458533#%E4%BD%93%E3%81%AE%E6%A7%8B%E9%80%A0)
図100 イカの体
地方独立行政法人 青森県産業技術センター
(https://www.aomori-itc.or.jp/public/~aosui/squid/ika2.html)
図101 ホメオティック突然変異
トライイット「5分でわかる!ホメオティック突然変異」
(https://www.try-it.jp/chapters-15244/sections-15293/lessons-15302/point-3)
図102 カンブリアンモンスター・アノマロカリス類
書籍「カンブリアンモンスター図鑑(秀和システム2015年)」に掲載された生物イラストレーシ
ョン
(https://www.behance.net/gallery/71364679/Cambrian-monster-picture-book)
図103 先カンブリア紀/カンブリア紀境界における海洋の無機炭素同位体比変動
Joachim E. Amthor; John P. Grotzinger; Stefan Schröder; Samuel A. Bowring; Jahandar
Ramezani; Mark W. Martin; Albert Matter, Geology (2003) 31 (5),431–434.
図104 ネオアサフス・コワレフスキー
フリー百科事典『ウィキペディア(Wikipedia)』「ネオアサフス・コワレフスキー」(https://ja.wikipedia.org/wiki/%E3%83%8D%E3%82%AA%E3%82%A2%E3%82%B5%E3%83%95%E3%82%B9%E3%83%BB%E3%82%B3%E3%83%AF%E3%83%AC%E3%83%95%E3%82%B9%E3%82%AD%E3%83%BC)
図105 地質年代と年代層序の区分
フリー百科事典『ウィキペディア(Wikipedia)』「地質時代」(https://ja.wikipedia.org/wiki/%E5%9C%B0%E8%B3%AA%E6%99%82%E4%BB%A3)
図106 後期カンブリア紀の正の炭素同位体変動
ResearchGate、Fig 1 – uploaded by Gong Soo Chung
(https://www.researchgate.net/figure/The-SPICE-Steptoean-Positive-Carbon-Isotope-Excursion-in-Laurentia-an-increase-in-d-13_fig1_296688523)
