
第2章 生命の起源
1.生命の起源におけるエネルギー
『代謝経路』(注27)は有機物の炭素が流れるネットワークである。
(注27)細胞内で進行する連鎖的な化学反応のこと。それぞれの経路において、元となる化学物質は一連の化学反応によって変化し、酵素がこれらの反応を触媒する。その適切な機能を果たすためには、酵素はしばしばミネラル、ビタミン、あるいはその他の補因子を必要とする。
「代謝経路」においては、連続した反応の触媒として作用する一連の酵素が存在する。それぞれの酵素は、前の酵素によって生成された物質に対して作用する。これにより、有機物の炭素の流れに制約が課される。分子が経路に参入すると、連続的に化学修飾を受け、新たな分子として経路を出る。これらの反応は繰り返し確実に行われ、同一の前駆体が反応に参入し、同一の生成物が出る。これら様々な代謝経路を通じて、細胞はネットワークを形成し、そこでの物質流れは常に制約を受けつつも、最大化が図られる。これらの緻密な制御によって、細胞は、無制限の流れよりもはるかに少ない炭素とエネルギーで成長することが可能となる。各ステップにおいてエネルギーを無駄にすることなく、酵素は生化学的メカニズムに対し正しい進行方向を強制する。細胞内では、エネルギー観点から言えば、酵素の役割は反応を加速することではなく、むしろその力の導入によって出力を最大化することである。
では、生命が起源を迎える段階で、酵素がまだ存在しなかった時にはどのような状態だったのだろうか。その時の流れの制約は必然的に少なかったはずである。有機分子をより多く生成し、倍加し、最終的には複製するためには、現代の細胞よりも多くのエネルギーや炭素が必要だったに違いない。現代の細胞は必要なエネルギーをできるだけ最小化している一方で、
まだ標準的なエネルギー通貨であるATPを大量に消費している。水素と二酸化炭素の反応によって育つ最高に単純な細胞でも、呼吸によって、新たに作り出すバイオマス(生物体量)のおよそ40倍もの老廃物を生み出す。つまり、新たなバイオマスを1グラム作るたびに、この産生を支えるエネルギーを放出する反応で、少なくとも40グラムの老廃物が生まれるはずなのである。
生命は、エネルギーを放出する主反応の副反応なのである(注28)。40億年の進化で洗練を経た現代でも、それは変わらない。現代の細胞が有機物の40倍の老廃物を生み出すとしたら、酵素を持たなかった最初の原始的な細胞がどれほどの老廃物を生み出していただろうか。酵素は化学反応を何百万倍も加速する。そんな酵素をなくすと、処理能力を同じぐらいの倍率、100万倍単位で増やさなければ、同じことができないだろう。
(注28)呼吸による有機物の分解で放出されたエネルギーを生命活動に直接利用することはできない。呼吸で放出されたエネルギーは、一時的に『ATP』という物質に蓄えられる。全ての生物が共通に用いるATPは、そのエネルギーが運動、生体物質の合成、発光などのさまざまな生命活動に使われるので、「エネルギー通貨」ともいわれる。ATPはアデノシンという物質にリン酸が3つ結合した『アデノシン三リン酸』の略称で、リン酸どうしの結合は「高エネルギーリン酸結合」と呼ばれる。呼吸により有機物から放出されたエネルギーで『ADP(アデノシン二リン酸)』と『リン酸』が結合してATPが合成される。ATPがADPとリン酸に分解されるとき、比較的大きなエネルギーが放出され、このエネルギーを用いて生命活動が営まれている。
したがって、最初の細胞は、1グラムの細胞を生成するのに、40トン(40グラムの100万倍)の廃棄物-すなわち、超大型トラック一台分-を生み出していた可能性がある。エネルギーの流れという観点から見ると、これは氾濫した河川というよりも、むしろ津波に近い。
- この純粋なエネルギー需要の規模は、生命の起源のあらゆる側面に影響を及ぼすはずであるが、これまでそれが明確に考慮されることはほとんどなかった。
実験科学の分野で、生命の起源の領域は1953年の有名な『ミラー-ユーリーの実験』にまで遡り、その結果はワトソンとクリックによる『二重らせん』の論文と同じ年に公表された。どちらの論文も、それ以来ずっとこの領域を支配し続けている。
ミラー-ユーリーの実験は、『原始スープ』の概念を強化し、その領域の視野を狭める効果を持った。一方で、クリックとワトソンはDNAと情報が支配する時代の到来を告げた。しかし、その結果、他の要素、特にエネルギーの重要性に対する認識が阻害されてしまったのである。
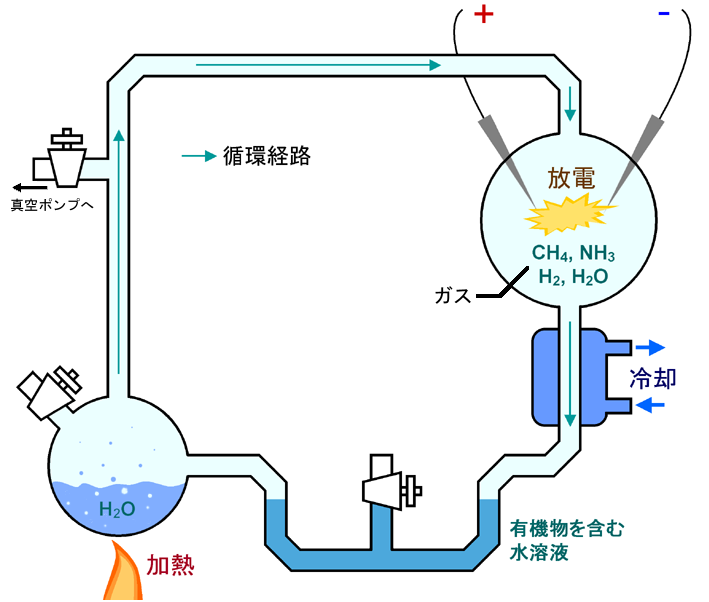
1953年、スタンリー・ミラーは、ノーベル賞受賞ハロルド・ユーリーの研究室で、若く熱意あふれる博士課程の学生だった。みずからの印象的な実験で、ミラーは木星の大気を模した還元性の(電子の豊富な)ガスの混合物を入れたフラスコ内で、稲妻をシミュレートした放電を行った。当時、木星の大気は初期の地球の大気を反映していると考えられていた。どちらも水素とメタンとアンモニアが豊富にあると推定されていた。驚くべきことに、ミラーは多くのアミノ酸の合成に成功した。アミノ酸はタンパク質の構成要素であり、生命の基本となる要素である。その結果、生命の起源は突如として容易に理解可能なものとなった。1950年代の始め、この実験には、ワトソンとクリックが見つけた構造よりもずっと大きな注目を浴びた。DNAの構造は初めて明らかにされたものの、当初はそれほど大きな話題にはならなかった。一方ミラーは、1953年に『タイム』誌の表紙を飾った。彼の成果は、生命の起源に関するはっきりした仮説を初めて検証したものだからという意味で画期的だったからである。
稲妻が還元性のガスの大気を通過して、細胞の構成要素ができたという仮説である。生命が存在しない状態では、これらの前駆体が海に集積し、やがて海は有機分子の濃厚なスープ、つまり『原始スープ』となったというストーリーである。
ワトソンとクリックが1953年に起こした騒ぎはそれより小さかったとしても、DNAの魅力はそれ以来ずっと生物学者をとりこにしてきた。
多くの人にとって、生命とは要するにDNAにコピーされた情報そのものである。したがって、生命の起源とは、自然選択による進化が不可能となるような情報の起源である。つまり、情報の起源は複製の起源ともなる。最初に自身のコピーを作る分子―『複製子』(注29)―が生じたプロセスに還元できるのである。DNA自体は非常に複雑で、最初の複製子だったとは考えにくい。しかし、より単純で反応性の高い前駆体であるRNAならばその条件を満たす。
(注29)『自己複製』とは、何らかの事物がそれ自身の複製を作る過程である。細胞は適当な条件が整うと、細胞分裂による複製を行う。細胞分裂において、DNAが複製され、生殖に際してはそれが子に転送される。ウイルスも複製されるが、細胞に感染して細胞の持つ生殖機構に指令を出すことでのみ複製可能である。
RNA(リボ核酸)は今日なお、DNAとタンパク質を結びつける重要な媒体であり、タンパク質の合成においてひな型と触媒の両方の役割を担う。RNAは、DNAのようにひな型と、タンパク質のように触媒の両方になりうるため、原初の『RNAワールド』においてはタンパク質とDNAの両方にとって、理論上はより単純な原型となる可能性がある。しかし、『ヌクレオチド』という構成要素はどこで生まれ、どのように鎖状に結合してRNAになったのだろうか。もちろん、答えは「原始スープ」であるとされる。RNAの形成とスープの間に必然的な関係があるわけではないが、それでもスープは最も単純な仮説であり、熱力学や地球化学などの込み入った細部に悩まされずに済む。
そうしたすべてを脇に置けば、遺伝子研究者は重要な話を進めることができる。したがって、
過去60年の生命起源研究に通底する核心的な思想があったとすれば、それは、
- 原始スープがRNAワールドを生み出し、そこでこの単純な複製子が次第に進化を遂げて複雑になり、代謝をコードして、最終的には今日私たちの知るDNAとタンパク質と細胞の世界を生み出した
というものなのだ。この観点から、生命はボトムアップ式の情報と見なせる。
- ここで見落とされているのは、『エネルギー』という要素である。
もちろん、エネルギーは原始スープにも関与している。稲妻が閃くからである。しかし、稲妻1回にはそれほど多くの電子は含まれていない。それに代わるもっと優れたエネルギー源は紫外線放射であり、これにより、メタンや窒素などの大気中の混合ガスから、シアン化物(およびシアナミドなどの誘導体)のような反応前駆体を作り出せる。紫外線は地球にもほかの惑星にも絶えず降り注いでいる。流れ込む紫外線は、オゾン層がなかった上に、太陽が若いと電磁スペクトルがさらに強烈なので、今より強かったはずである。聡明な有機化学者ジョン・サザーランドは、紫外線放射とシアン化物を用い、いわゆる「妥当な原初の条件」のもとで、活性化したヌクレオチドの合成に成功さえしている。
しかしながら、地球上にはシアン化物を炭素源として利用している生命や、紫外線をエネルギー源として利用している生命は存在しない。それどころか、シアン化物や紫外線は危険な殺し屋として認識されている。紫外線は現代の高等生物に対しても極めて有害である。これは、有機分子の形成を促進するよりも効果的にそれを分解するからだ。生命で海を満たすよりも、生命を焼き尽くす可能性の方が遥かに高い。地球であろうと、他の場所であろうと、直接的なエネルギー源となるとは考えにくい。
ヘラクレイトスが「同じ川には二度と入れない」と教え、つまり、見た目は同じでも毎回川が変わっているという教えを残しているが、それと同じように、生命も形を保ちながら中身を絶えず更新している。生物の細胞は形を保ちながらも、その構成要素は絶えず代謝回転(全体としての量は変わらないが代謝により更新される状態)によって交換されている。他の形は存在しえなかったのかもしれない。
- 複製子の登場以前、生命誕生の際には必然的にそうだったに違いないが、「構造」を規定する情報がなくても「構造」は存在した。
- ただし、そのためには継続的な『エネルギーの流れ』を必要であった。この「エネルギーの流れ」により物質には「自己組織化」が促されるからである。
- 具体的には、ロシア生まれのベルギーの大物理学者イリヤ・プリゴジンが『散逸構造』(注30)と呼んだ現象で、これは私たちにとってもなじみ深いものである。
(注30)『散逸構造』とは、熱力学的に平衡でない状態にある「開放系構造」を指す。すなわち、エネルギーが散逸していく流れの中に自己組織化の基が生まれる定常的な構造である。イリヤ・プリゴジンが提唱し、ノーベル賞を受賞した概念で、「定常開放系」、「非平衡開放系」などとも表現される。
もう少し具体的に説明するために、ここで簡単に、『散逸』という概念について考えてみる。「エネルギー」にはいろいろな形があるが、代表的なものとして「運動エネルギー」を取りあげてみると、たとえば、床を転がるボールは運動エネルギーを持っている。しかし、転がるうちに徐々に速度が遅くなり、最終的には止まる。これは、ボールの運動エネルギーが床との摩擦によって摩擦熱に変化してしまうためである。つまり、運動エネルギーが熱エネルギーに変化したのである。しかし、この逆の現象は起こらない。止まっているボールが床から熱を集めて突然転がり始めることはない。このように、エネルギーの変換には向きがある。運動エネルギーだけでなく、いろいろなエネルギーが熱エネルギーに変化するが、その逆は起こらない。このように一方向性を持ち、逆向きの過程が起こらない現象を『不可逆過程』と呼ぶ。そして、いろいろなエネルギーが熱エネルギーに変化する不可逆過程のことを、『散逸』と表現する。散逸構造は、岩石のようにそれ自体で安定した自らの構造を保っているような構造とは異なり、例えば「潮」という運動エネルギーが流れ込むことによって生じる内海の渦潮のように、一定の入力のあるときにだけその構造が維持され続けるようなものを指す。
沸騰するやかんの中の対流や、シンクの穴に水が渦を巻いて流れ込む様子を思い浮かべてみてもよい。情報は必要ない。やかんの場合は熱だけ、シンクの穴の場合は角運動量だけあればよいのである。
散逸構造は、エネルギーや物質の流れによって形成されるものである。
ハリケーン、台風、渦潮は、自然界で観察される散逸構造の典型例である。さらに大きなスケールでは、海洋や大気において、赤道と極で太陽から受けるエネルギーの流れの差が大規模な散逸構造を生み出す。メキシコ湾流のような定常的な海流や、「吠える40度」や北大西洋の「ジェット気流」などの風は、情報によって制約されることなく、それらを維持するエネルギーの流れと同様に、安定的に継続している。木星の「大赤斑」は巨大な嵐であり、地球数個分の規模の高気圧エリアとなっており、少なくとも数百年にわたって維持されてきた。やかんの中の対流セルも、電熱線に流れる電流が水を沸騰させ、蒸気を発生し続ける限り、維持される。これらすべての散逸構造は、エネルギーの継続的な流れを必要とする。
さらに一般化すると、これらは持続的な非平衡状態が結果として可視化されたものと言える。その状況下では、エネルギーの流れが、最終的に(恒星の場合は何十億年後に)平衡状態に達し、構造が崩壊するまで、構造を維持し続ける。つまり、
- 持続的で一定の物理的構造が、エネルギーの流れによって生み出せるのである。
全ての生物は、非平衡な状況下で維持されている。私たち自身も散逸構造の一部である。呼吸による絶え間ない反応は、細胞が炭素を固定し、成長し、反応中間体を形成し、その構成要素を組み合わせて炭水化物やRNA、DNA、タンパク質などの長鎖ポリマーを合成し、低エントロピー状態を周囲のエントロピーの増大によって維持するための自由エネルギーを供給する。
- 遺伝子などの情報がなくても、細胞膜やポリペプチドなどの細胞構造は、反応前駆体-活性化したアミノ酸やヌクレオチド、脂肪酸-が絶えず供給され、必要な構成要素を供給する継続的なエネルギーの流れが存在する限り、自然に形成される。
- 細胞構造は、エネルギーと物質の流れによって必然的に発生する。部品が交換されても、構造は安定しており、流れが続く限り維持される。
- この『エネルギーと物質の継続的な流れ』こそ、原始スープに欠けているものなのである。原始スープには、私たちが『細胞』と呼ぶ散逸構造を形成する促進因子が存在しない。代謝を誘導して促す酵素がなくてもこうした細胞を成長・分裂させ、細胞の活動を維持するための要素が欠けているのである。
しかし、そもそも「最初の原始細胞を形成するための環境」は本当に存在したのであろうか。多くの疑問が持たれるかもしれないが、そのような環境が存在した―具体的には後述する『アルカリ熱水噴出孔』がそれに該当する―という事実を以下で説明する。しかし、その環境を探求する前に、『細胞』と呼ばれる散逸構造の形成を促すために具体的に何が必要なのかを考察することが重要である。
細胞の作り方
細胞を作るのには何が要るだろうか。
実は、地球上の生体細胞は「6つの基本的特質」を共通して持っている。それらを上げると以下のとおりである。
- 新たな有機物の合成のために反応性の高い炭素が継続的に供給されること
- 代謝の生化学的メカニズム-新たなタンパク質やDNAなどの形成-を働かせる自由エネルギーが供給されること
- こうした代謝反応を加速し誘導する触媒の存在
- 熱力学第二法則(注31)に従い、化学反応を正しい方向に進行させるために、廃物を排出すること
(注31)『トムソンの原理(ケルビンの法則)』とも呼ばれ、「一つの熱源から熱を受け取り、そのすべてを仕事に変換することは不可能である」という原理である。つまり、「熱の全てを運動として取り出すことはできない」ということ。例えば、自動車のエンジンで説明すると、エンジンがガソリンを燃やして生じる「熱」を「ピストンの動き」に変換するが、100%ピストン運動に変えることはできない。一部が必ず「排熱」として排出される。すなわち、高温の熱エネルギーから仕事を取り出すには、低温の熱源に熱を捨てなければならないのである。言い換えると、「熱を全て仕事に変える熱機関は存在しない」というのが 「トムソンの原理(ケルビンの法則)」である。
- 区画化-内側と外側を隔てる細胞状の構造
- 遺伝物質-具体的な形状や機能を規定するRNAやDNA、あるいはそれと同等のもの
一見して、6つの因子は全部強く依存し合っていて、一番初めからそうでなければならないこともほぼ確実だとわかる。有機物の炭素が継続的に供給されることは、成長や複製などのすべてにとって、明らかに肝要だ。単純なレベルでは、RNAワールドさえRNA分子の複製を必要とする。RNAは『ヌクレオチド』という構成要素が連なった鎖で、個々のヌクレオチドはどこかからやってきたはずの有機分子だ。代謝と複製のどちらが先に現れたかについては、生命起源の研究者の間で古くから意見の相違があり結論の見えない論争だが、いずれにせよ、複製とは倍加する過程であるため、構成要素を幾何級数的に消費する。同様の速度でこれらの構成要素が補充されなければ、複製はすぐに停止する。したがって、
- なんらかの初期の地球環境が、複製の誕生に必要な有機物の構成要素、活性化したヌクレオチドを提供できたのだと仮定するのは理に適っている。
- 還元性大気中の放電から、小惑星での宇宙空間の化学反応や、高圧の爆発反応装置に至るまで、異なる条件できわめてよく似た一連の有機物ができるという傾向は、ある種の分子-いくつかのヌクレオチドも含まれるに違いない-が熱力学的に好まれることを示唆している。
従って、
- 有機物の複製子が形成されるには、同じ環境のなかで有機物の炭素を継続的に供給する必要がある。
では、エネルギーについてはどうだろうか。これもまた同じ環境の中で必要となる。アミノ酸やヌクレオチドといった個々の構成要素を結合して長鎖のポリマー(タンパク質やRNA)を作るためには、まず構成要素を活性化しなければならない。その次に必要となるのはエネルギー源であり、それはATPか、それに類似した何かである。おそらく非常によく似たものであろう。
- 40億年前の地球がそうだったように、全面が水で覆われた世界では、エネルギー源は特異なタイプであり、長鎖分子の重合を促進する必要がある。そのためには、新たな結合が形成されるたびに水分子を1つ取り除く、すなわち『脱水反応』が必要となる。
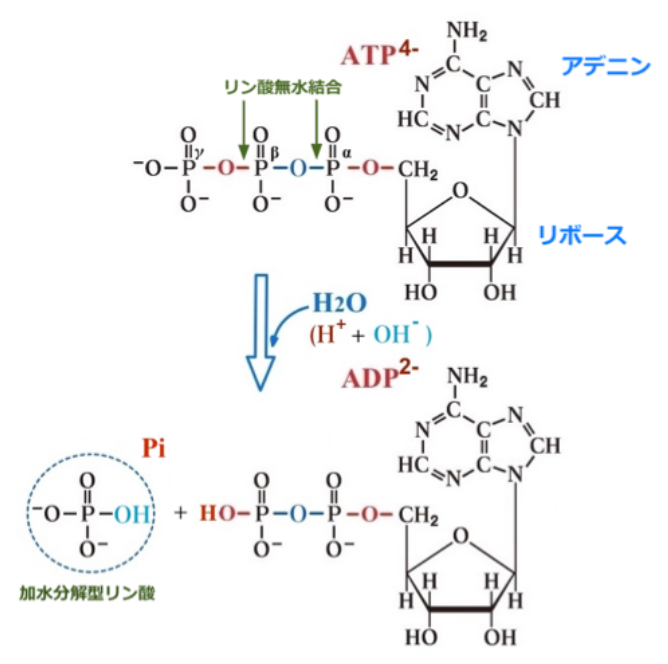
溶液中で分子を脱水するという課題は、水中で濡れた布から水を絞り出そうとすることに少し似ている(注32)。しかし、地球上の生命は水中でも問題なく活動している。すべての生体細胞は、1秒間に何千回もの脱水反応を達成している。我々は、この反応をATPの分解と組み合わせることで達成している。一度の分解で1つの水分子が取り除かれる。
(注32)ATPの合成と分解の化学反応はタンパク質によって触媒されており、ATPの合成と分解は 水中では容易には起こらないことが知られている。このことよりATPの三次元構造は、タンパク質と相互作用している場合と水中では大きく異なっていることが予想される。
- 『脱水反応』と『加水分解反応』の組み合わせは、基本的には水の移動に過ぎないが、それと共にATPの結合に閉じ込められていたエネルギーの一部が解放される(注33)。このように考えると、問題は大いに単純化される。
(注33)ATPが蛋白質に結合すると上図に示す加水分解反応「ATP+H20→ADP+Pi」が起こり、その時に放出される30.5kJ/mol (=7.3kcal/mol) ものエネルギーが様々な蛋白質の機能発揮のエネルギー源となる。
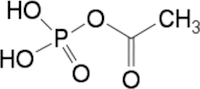
- 重要なのは、ATPか、それと同等だがより単純な『アセチルリン酸』などが、絶えず供給されることである。要するに、水中での複製には、同じ環境の中で、有機物の炭素とATPのようなものが継続的に惜しげなく供給される必要がある
ということである。これで、6つの因子のうち3つ、つまり「複製」、「炭素」、「エネルギー」について考察した。
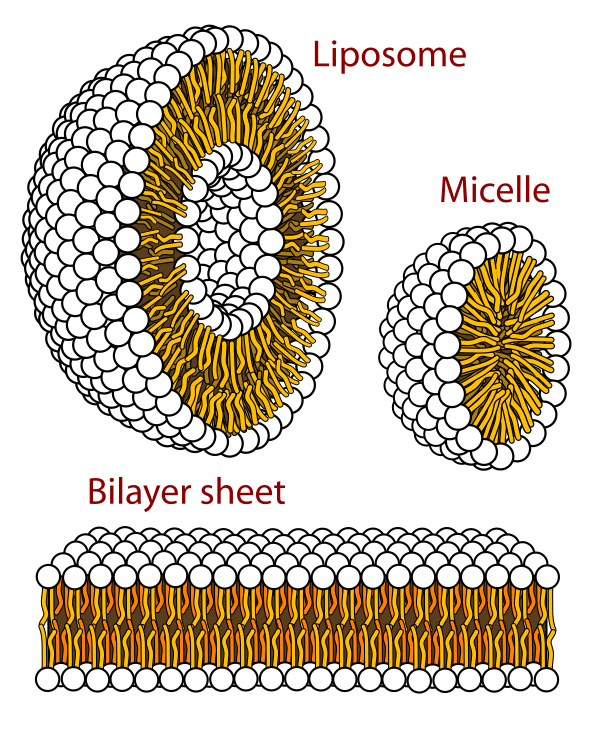
次に、「細胞形成への区画化」について考えてみよう。これも濃度の問題である。
生体膜は『脂質』(<物語り その3>図26参照)でできており、脂質そのものはまた『脂肪酸』などで構成されている。ある閾値を超える濃度になると、脂肪酸は自然に細胞状の小袋を形成する(図42参照)。そして新たな脂肪酸が継続的に供給されれば成長し分裂することが可能となる。ここでも、新たな脂肪酸の合成を進めるためには、有機物の炭素とエネルギーの両方が継続的に供給される必要がある。
- 脂肪酸やヌクレオチドといったものが散逸する速度を上回って蓄積するためには、何らかの集約が必要である。
- 物理的に集めたり天然の区画化を行ったりして、局所的に濃度を高め、より大きな規模に構造を形成できるようにするのである。
このような条件が満たされれば、小袋の形成はもはや魔法ではない。これが物理的に最も安定な状態であるからだ。前章「生命」の「エネルギー、エントロピー、構造」の節で見たとおり、結果的に全体のエントロピーは増大するのである。
反応性のある成分が継続的に供給されると、「表面積対体積」の制約により、単純な小袋は自然と成長して分裂するのである。
球状の小袋-単純な『細胞』-の中に、さまざまな有機分子が収められているとしよう。小袋は新たな素材-膜には脂質、細胞内にはそれ以外の有機物-を取り込むことで成長する。サイズを倍、すなわち、膜の表面積と細胞内の有機物を倍とした場合を考えてみよう。表面積が倍になると、体積は倍よりも大きくなる。これは表面積が半径の2乗に比例し、一方で体積は3乗に比例して増大するためである。しかし、細胞内の有機物の増大は倍であるので、有機物の量が膜の表面積より速く増大しなければ、小袋は縮んでダンベル状になり、新たな二つの小袋を形成する途中の状態となる。つまり、
- 算術的な成長が、ただ大きくなるだけでなく、分裂や倍増へ導く不安定性をもたらすのである。やがて成長する球体が小さな泡に分裂するのも時間の問題にすぎない。
- そのため、反応性の高い炭素という前駆体が継続的に流れ込むと、必然的に原始的な細胞の形成のみならず初歩的な形態の細胞分裂も起こることになる。
「表面積対体積」の比率問題は、細胞のサイズに制約を与え、反応物質の供給と老廃物の除去の問題を引き起こすのである。
細胞の場合、新たなタンパク質が形成される速度は、反応する前駆体(活性化したアミノ酸)が運ばれる速度と老廃物(メタン、水、CO2、エタノール-エネルギーを放出する反応が何であれ)が除去される速度に依存する。こうした老廃物が細胞から物理的に取り除かれないと、反応が続かなくなる。
- 「老廃物の除去」という問題は、反応物質と老廃物が一緒に漬け込まれた原始スープの概念を阻害する、別の根本的な障害となる。新たな化学反応を進行させる原動力が存在しないのである。
さらに、細胞が大きくなるほど、その状態はスープに近づく。細胞の体積は表面積より速く増大するため、細胞が大きくなるほど、膜を超えて新たな炭素を運び込む速度や老廃物を除去する速度が相対的に低下する。大西洋サイズの細胞、あるいはサッカーボールのサイズの細胞でさえ、適切に機能することはなく、それは単なるスープである。
- 生命の起源では、炭素の供給と老廃物の除去の速度から考えると、細胞の体積は小さくあることが求められる。何らかの物理的な誘導ー前駆体を運び入れ、老廃物を運び出す継続的な自然の流れーも必要となるだろう。
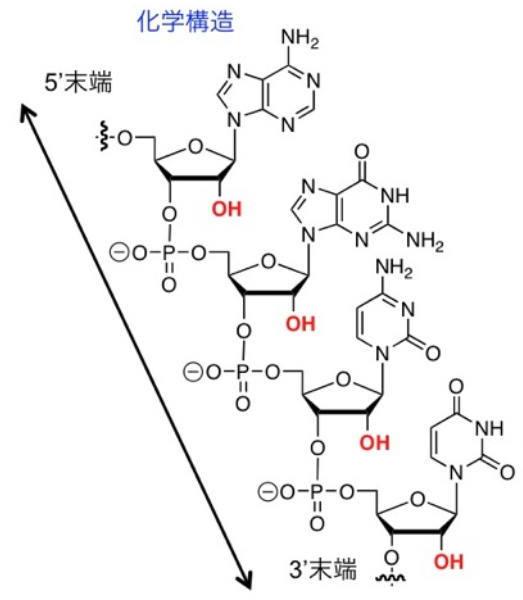
次に、「触媒」について考えてみる。現代の生命はタンパク質-酵素-を用いているが、RNAにも一定の触媒能力が存在する。問題となるのは、RNAが既に複雑なポリマーであるという事実である。RNAは多くのヌクレオチドという構成要素から成り立ち、個々のヌクレオチドを合成し、活性化して連結し、長い鎖を形成しなければならない。したがって、RNAが触媒として機能する前に、これらの過程が必要となる。どのようなプロセスがRNAを生成したにせよ、それはより合成しやすい有機分子、特にアミノ酸や脂肪酸の生成をも促進したと考えられる。したがって、初期の「RNAワールド」には、多種多様な小さな有機分子が混在していたと推測される。
- たとえRNAが複製やタンパク質合成の起源で重要な役割を果たしたとしても、RNAが単独で代謝を生成したとは考えにくい。
では、何が生化学的メカニズムの発端で触媒となったのだろうか。
可能性として考えられるのは、『金属硫化物』(とくに鉄、ニッケル、モリブデンのもの)など、『無機の錯体』である。これらは今も、いくつかの古い、広く保存されているタンパク質に『補因子』(酵素の触媒活性に必要なタンパク質以外の化学物質)として存在している。
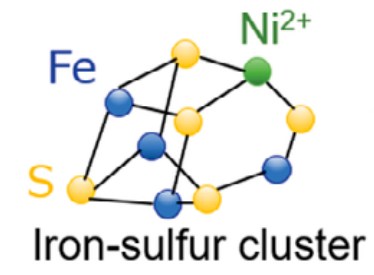
- タンパク質は触媒として認識されがちであるが、それは実際にはどのようにしても起こる反応を加速させるだけで、補因子が反応の本質を決定している。タンパク質という環境がなければ、補因子はあまり効果的な触媒とはならず、特異性も低い。しかし、何もない状態よりははるかに効果的である。その効果は、処理能力に依存する。初期の無機触媒は、有機物への酸素とエネルギーの供給を始めたのみで、その流れは津波でなく小川程度で良いとしたのである。
- 単純な有機物、特にアミノ酸やヌクレオチド(図39参照)には、それ自体に触媒作用がある程度存在する。アセチルリン酸(図41参照)の存在下では、アミノ酸は連結し、短いポリペプチド-つまりアミノ酸の小さな鎖-を形成することが可能である。そのようなポリペプチドの安定性は部分的に他の分子との相互作用によって影響を受ける。脂肪酸と結合する疎水性のアミノ酸やポリペプチドは比較的長期間保持され、FeSなどの無機クラスターと自然に結合すると鉱物の触媒特性を強化する可能性がある。このため、物理的に保持されることにより「選択される」可能性がある(注34)。
(注34)「アミノ酸」は、分子内にアミノ基(-NH2)とカルボキシル基(-COOH)を持つ化合物の総称である。アミノ基とカルボキシル基が結合する炭素の位置により、α、β、γ、δ、εなどのアミノ酸が存在するが、タンパク質を構成するアミノ酸は全てα-アミノ酸である。また、「ペプチド」は、一般的に、2~50個程度のアミノ酸がペプチド結合したものを指す。2個のアミノ酸が結合したものは「ジペプチド」、3個では「トリペプチド」と呼ばれる。アミノ酸の数が2~20個程度のものは「オリゴペプチド」、さらに多くのアミノ酸が結合すると「ポリペプチド」と称される。
次に、有機合成を促進する鉱物の触媒を考察する。
合成された有機物の中には、鉱物の触媒と結びつきながら生存し、触媒特性を強化する(または少なくとも変化させる)ものも存在する。そのようなシステムは、原理的に、より複雑な有機化合物をより豊かに生成する可能性があるのではないか。
そこで問題となるのは、どのようにして細胞を一から作り出すかである。
反応性の高い炭素と利用可能な化学エネルギーは、持続的かつ大量に供給される必要がある。この過程で、流れの一部が初期の触媒を通過して新たな有機物に変換される。
- この継続的な流れには、老廃物の流出を妨げずに、脂肪酸やアミノ酸やヌクレオチドなどの有機物を高濃度にため込めるような、なんらかのやり方で制約を加えなければならない。
- そのような流れの集中は、自然の誘導や区画化によって達成可能で、これは水車の流れの誘導と同様の効果をもたらす。酵素なしに任意の流れの力を増すので、必要な炭素やエネルギーの総量が減らせるのだ。
- 新たな有機物の合成が外界への損失を上回り、濃縮されると、有機物は細胞状の小袋やRNAやタンパク質(ポリペプチド)などの構造に自己組織化する。
ただし、これはまだ細胞の始まりにすぎない。必要だが、決して十分ではない。しかし、細かいことはとりあえずさておいて、この一点だけに目を向けよう。
- 大量の炭素とエネルギーの流れが無機の触媒を通過するように物理的に誘導されなければ、細胞の進化は不可能である。これが、宇宙のどこにおいても必要な条件である。
- 前の節で述べた炭素の化学反応の必要性を考えると、熱力学的には、炭素とエネルギーが自然の触媒を通じて持続的に流れることが要求される。
したがって、生命の起源として提唱されてきたほぼすべての環境―「温かい水たまり(ダーウィン)」、「原始スープ(ユーリー–ミラーの実験)」、「微小な孔のある軽石」、「海辺」、「パンスペルミア(他の天体で発生した微生物の胞子説)」など―は、その対象から排除される。
ところが
『熱水孔』はその例外である。むしろ、これを積極的に採用する。なぜなら、「熱水孔」は、まさに我々が求めているタイプの散逸構造、すなわち、持続的な流れを持ち、平衡状態からは遠い「電気化学反応装置」であるからだ。
熱水孔は流通反応装置
熱水孔は、一種の流通反応装置である。だが、『熱水孔』(注35)に対する注意も必要である。
(注35)『熱水孔(熱水噴出孔)』は、地熱で熱せられた水が噴出する大地の亀裂である。広義の熱水噴出孔としては温泉・噴気孔・間欠泉も含まれるが、狭義にはこれらの陸上にあるものではなく、海底環境、特に深海の「熱水噴出孔(深海熱水噴出孔)」のことを指す。熱水噴出孔の英語表記やその構造物から、「ベント(vent)」や「チムニー(chimney)」と呼ばれることもある。熱水噴出孔の大半は、火山活動が活発なところ(発散的プレート境界、海盆、ホットスポット)から発見されている。吹き出す熱水は数百度にも達することがあり、溶存成分として重金属や硫化水素を豊富に含むものも知られている。海底から噴出する熱水に含まれる金属などが析出・沈殿してチムニーと呼ばれる構造物が形成される場合がある。熱水の溶存成分によってはチムニーから黒色や白色の煙が吹き出しているように見えるため、一部の熱水噴出孔は『ブラックスモーカー』や『ホワイトスモーカー』と呼称される場合もある。
現在の熱水孔は太陽から隔絶されているわけではない。ここに生息する生物は、スモーカーから出る硫化水素ガスを酸化する細菌との共生関係に依存している。これが非平衡の主要な原因である。硫化水素(H2S)は還元性のガスで、酸素と反応してエネルギーを放出する。先の章で触れた呼吸のメカニズムを思い出すと、細菌は呼吸によりH2Sを電子供与体、酸素を電子受容体として用い、ATP合成を促進する。しかし酸素は光合成の副産物で、酸素発生型光合成が誕生する前の初期の地球には存在しなかった。したがって、『ブラックスモーカー』の熱水孔の周囲で生命が豊富に存在する事実は、間接的にではあるが完全に太陽のおかげである。このような熱水孔は40億年前には全く異なった存在であったと推測される。
では、酸素がなければ、どのような可能性が残るだろうか。
実は、ブラックスモーカーは、『中央海嶺(注36)の拡大中心』(マントルからプレートが上昇して広がるところ)などの火山活動が活発な場所で、海水とマグマが直接相互作用して生じる。
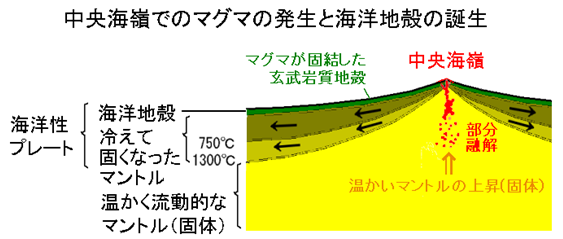
(注36)海洋底には、高温のマントル物質が浅いところまで上昇してマグマが発生し、海底火山の活動が盛んな山脈がある。このようなところを『海嶺(かいれい)』と呼ぶ。その海嶺のうち、海洋プレートが生成され海洋底が拡大する大規模な海底山脈を『中央海嶺』と呼ぶ。代表的な中央海嶺である『大西洋中央海嶺』は大西洋の中央部に南北に連なる大きな裂け目で、東西方向に引っ張られてプレートが拡大している。
海底に浸透した水が、それほど深くない場所にあるマグマのたまり場に達し、そこで一気に数百度まで温度が上昇し、マグマに溶存する金属や硫化物を取り込んで強い酸になる。過熱した水は爆発的な力で上方へ突き上げられ、急速に冷える。黄鉄鉱などの硫化鉄の微粒子が即座に析出し、これが黒い煙(ブラックスモーク)を生成し、「ブラックスモーカー」の名を与える。
その構造の大半は40億年前も同じだったと考えられる。しかし、そこで生成されるFeS鉱物が優れた触媒であったとしても、熱水孔の250~400℃の温度で最も安定した炭素化合物はCO2であるため、有機合成は起こらないと判明している。また、ブラックスモーカーは非常に不安定で、せいぜい数十年で成長し崩れるため、生命を「生み出す」には時間が足りない。
ブラックスモーカーは真に平衡とはほど遠い散逸構造で、確かにスープの問題の一部を解決できるが、こうした火山性のシステムはひどく極端で不安定なので、生命の起源に必要な穏やかな炭素化学反応を育めない。
- ただ、ブラックスモーカーなどの熱水噴出孔は、マグマに由来する第一鉄(Fe2+)イオンやニッケル(Ni2+)イオンなど、触媒となる金属で初期の海を満たすという重要な役割を果たしたということは認識すべきである。
海に溶け込んだこうした金属の恩恵を受けるのは、『アルカリ熱水噴出孔』という別のタイプの熱水孔だった。この熱水孔でブラックスモーカーの問題がすべて解決できるのである。
- 『アルカリ熱水噴出孔』は火山のようなものではなく、ブラックスモーカーの華々しさや刺激を欠いてはいるが、電気化学的な『流通反応装置』(所定の温度、圧力に保たれた装置に原料を連続的に供給して反応を行わせ、連続的に抜き出す形の反応装置)としてはるかによく整えられた特性をほかに持っている。

実は、『アルカリ熱水孔』は、水とマグマとの相互作用ではなく、はるかに穏やかなプロセス、すなわち「固体岩石と水との化学反応」によって生み出されている。マントルに由来する岩石は、『カンラン石』(上部マントルの主要な構成鉱物で、フォルステライト(Mg2SiO4)とファヤライト(Fe2SiO4)が規則正しく混ざった固溶体)などの鉱物を豊富に含み、水と反応して含水鉱物の『蛇紋岩』を形成する。蛇紋岩は、蛇の鱗に似た美しい緑色の斑点模様を持つ。蛇紋岩を磨いたものは、緑の大理石のように、ニューヨークの国連ビルなど公共の建物で、一般に装飾石として利用されている。
この岩を形成する化学反応は、『蛇紋岩化作用』(注37)といういかつい名前を持っているが、要するにこれは、カンラン石が水と反応して蛇紋岩を形成するということであり、この反応の「廃棄物」が、生命の起源の鍵を握っていたのである。
(注37)「蛇紋岩化作用」の化学反応式は以下のとおりである。
3(Mg0.9Fe0.1)2SiO4+4.1H2O→1.5Mg3Si2O5(OH)4+0.9Mg(OH)2+0.2Fe3O4+0.2H2(aq)
カンラン石 水 蛇紋岩 ブルーサイト 磁鉄鉱 水和水素
蛇紋岩化作用は「カンラン石((Mg0.9Fe0.1)2SiO4)」と水が反応することで引き起こされ、その結果として、蛇紋岩(Mg3Si2O5(OH)4)、ブルーサイト(Mg(OH)2) [補注:「水滑石」とも呼ばれ、天然に鉱物として産出する水酸化マグネシウム]、磁鉄鉱(Fe3O4)、 水和水素(H2(aq))[補注:溶媒である水分子と結合した状態、または強い相互作用下にある現象]が生じる。蛇紋岩化作用に必要なカンラン石は上部マントルの主成分であるカンラン岩中に多量に含まれている。沈み込みプレート境界などで海水や海水を含んだ含水鉱物がプレートとともにマントルに引き込まれると、地下深部でカンラン石と水が反応し、蛇紋岩化作用が起きる。なお、磁鉄鉱(Fe3O4)は、赤鉄鉱とともに鉄の重要な鉱石鉱物で、第一鉄(Fe2+)と第二鉄(Fe3+)を1:2のモル比で含んでいる。
カンラン石には第一鉄とマグネシウムが豊富に存在する。第一鉄は、水によって酸化され、錆びた形態である第二鉄になる。この反応は熱を放出する発熱反応で、水酸化マグネシウムを含む温かいアルカリ流体に溶け込んでいた大量の水素ガスを発生させる。カンラン石は地球のマントルに豊富であるため、この反応は主に構造プレートの拡大中心付近の海底で起こる。新しく形成されたマントル岩石が海水に曝される場所である。
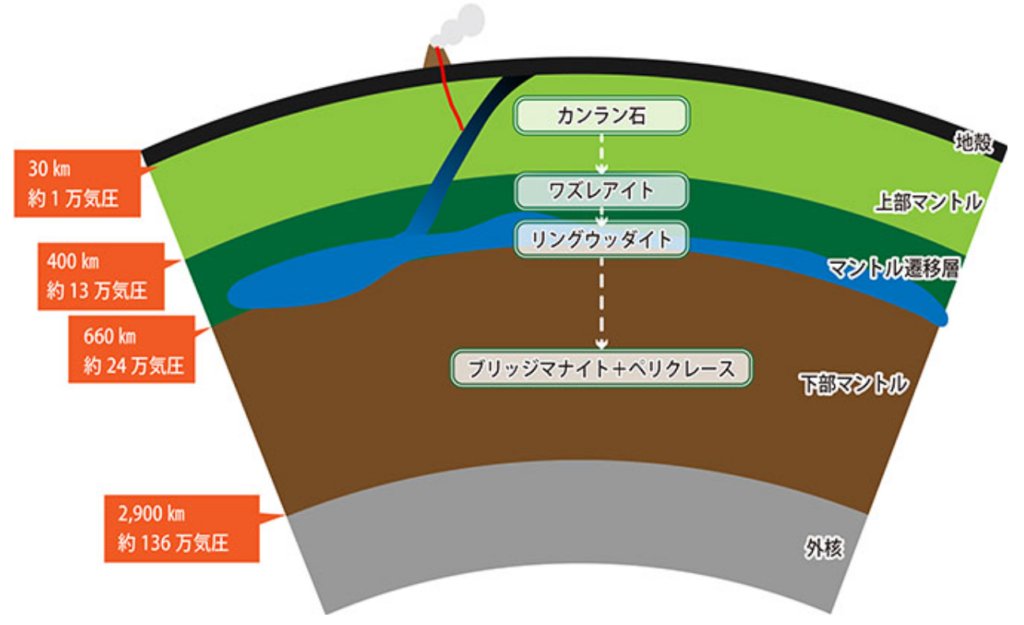
しかしながら、マントル岩石が直接水に曝されることは稀で、水は海底から侵入し、時に数キロメートルの深さまで達し、カンラン石と反応する。こうして生じた水素が豊富な温かいアルカリ流体は、下降する冷たい海水よりも軽く、海底に向けて上昇する。海底に達すると、その流体は冷却され、海水に溶け込んだ塩と反応し、大きな熱水孔を形成する。
ブラックスモーカーとは異なり、「アルカリ熱水孔」はマグマとは無関係であり、そのため拡大中心でのマグマだまりの直上ではなく、通常は数キロメートル離れた場所に存在する。温度は過熱状態ではなく、60~90℃の温熱状態である。その熱水孔は、海に直接煙のように噴出する大きな孔ではなく、相互に接続された迷路のような微細な孔が密集している。そして、酸性ではなく、強いアルカリ性を示す。

2000年12月に、20階建てのビルに相当する白い柱が林立するアルカリ熱水孔が初めて見つかり、『ロスト・シティー』(図48)と名づけられた(注38)。驚いたことに、ロスト・シティーは、生命の起源との関連性を1988年にはじめて『ネイチャー』の短いレター(注39)で示唆した革新的な地球化学者マイク・ラッセルの予測のほぼすべてと、大西洋中央海嶺から15キロメートルあまりというその場所に至るまで一致していたのである。
(注38)2000年12月、21世紀の到来を目前に控えて、大西洋中央海嶺から西へ15km程度の地点に位置するアトランティス岩体を調査した結果、全く新たなタイプの「熱水噴出孔」が見つかった。この噴出孔は20階建てのビルに匹敵する高さの白い柱が林立しており、『ロスト・シティー』と名付けられた。それまで知られていた「ブラックスモーカー」と呼ばれる熱水噴出孔は強酸性であり、その水温は400℃を超えていた。しかし、そのような過酷な環境にも関わらず、その周辺には1mを超えるチューブワームやカニといった生物が生息していた。一方、新たに発見された「ロスト・シティー」は強アルカリ性を示し、水温は90℃までで、生物が介在せずにメタンやプロパンといった有機物が生成され、エネルギー豊かな水素ガスが噴出していた。さらに、「ロスト・シティー」には太陽エネルギーと光合成による酸素を必要としない微生物が独自の生態系を築いていたのである。
(注39)M.J. Russell and A.J. Hall, Nature 336, 117 (1988), SCIENTIFIC CORRESPONDENCE
アルカリ性であることの重要性
『アルカリ熱水噴出孔』は、まさしく生命の起源に必要な条件を提供する。「炭素とエネルギーの大量の流れを、無機の触媒のもとを通過するように物理的に誘導し、有機物を高濃度にため込めるやり方で制約する」という条件である。
熱水流体は水素を豊富に溶解し、一方でメタン、アンモニア、硫化物などの還元性ガスの含有量は比較的少ない。ロスト・シティーを含む既知のアルカリ熱水孔は多数の細孔を持つ。中央に立つチムニーはないが、岩石そのものは鉱化したスポンジのように見え、薄い壁で隔てられた孔は相互に連絡しており、マイクロメートルからミリメートルオーダーの大きさを持つ、全体としての巨大な迷路を形成している。この迷路はアルカリ熱水流体により浸透されている。これらの流体はマグマにより過熱されていないため、その温度は有機分子の合成に適しており、流速の低下にも有利である。流体は急速に噴出されるわけではなく、触媒の表面をやさしく撫でるように進む。このような熱水孔は何千年も持続し、ロスト・シティーの場合は10万年以上にも及ぶのである。
この迷路状の細孔を通過する熱水の流れは、有機分子(アミノ酸、脂肪酸、ヌクレオチドなど)を「熱泳動」(注40)というプロセスにより、数千倍から数百万倍にまで劇的に濃縮する能力を有する。
(注40)温度勾配が存在する場所に微小粒子が存在する場合、高温側の気体から受ける運動量は低温側の気体から受ける運動量よりも大きくなる。これは微小粒子が高温側から低温側へ移動する力を受けることを意味し、この力を『熱泳動力』と呼ぶ。
- すべては運動エネルギーによって決まる。高温下では小さな分子が活発に動き、一定の自由度を持つ。しかし、熱水流体が混ざり合い冷却すると、有機分子の運動エネルギーが低下し、分子の動きの自由度が制限される。この結果、有機分子は細孔から離れにくくなり、低運動エネルギーの領域に集まる。
- 熱泳動の効果は分子のサイズに依存する。ヌクレオチドなどの大きな分子は、小さな分子よりも細孔内に留まりやすい。一方、メタンなどの小さな最終生成物は、熱水孔から失われやすい。
要するに、
多数の細孔を通過する熱水流体の流れは、安定した状態の条件を変化させるのではなく、安定状態に維持するような動的なプロセスを介して、有機物を積極的に濃縮する。さらに便利なことに、熱泳動は有機物同士の相互作用を促進し、熱水孔の細孔内で散逸構造の形成を進行させる。
- このような散逸構造は自然に脂肪酸を小袋にし、場合によってはアミノ酸やヌクレオチドを重合させてタンパク質やRNAを形成する可能性がある。
- このような相互作用が発生するかどうかは濃度による。どのようなプロセスで濃度を高めるにせよ、それは分子同士の化学的相互作用を促進するのである。
これは出来過ぎのように思えるかもしれないが、実際にその通りである。ロスト・シティーのアルカリ熱水噴出孔は、現在、多くは比較的普通の細菌や古細菌によって占められており、多様な生命体の生息地となっている。また、低濃度の有機物、例えばメタンや微量の炭化水素なども産生している。しかしながら、この熱水孔は現在では新たな生命体を創出してはおらず、熱泳動によって有機物が豊富な環境を形成してもいない。その理由は一部が、既にそこに住んでいる細菌が利用可能な資源を非常に効率よく消費しているからである。だが、それよりももっと根本的な理由も上げられる。
現在のブラックスモーカーが40億年前とは全く異なった特性を持っていたと同様に、アルカリ熱水噴出孔もその化学的性質が異なっていたに違いない。とはいえ、一部の特性は非常に類似していたと考えられる。例えば、
- 蛇紋岩化作用のプロセスそのものはどこも異なっていなかったはずだし、同じ温かく水素の豊富なアルカリ流体が海底へ向けて上昇していたと推察される。
- しかし、当時の海洋の化学的性質は現在とは大きく異なっており、それゆえにアルカリ熱水噴出孔の鉱物組成も変化していたことは自明である。現在、ロスト・シティーはほぼ炭酸塩(特にアラゴナイト(注41))で構成されているが、より最近に発見された他の類似した熱水噴出孔(アイスランド北部のストリィタンなど)は、主に粘土で形成されている。
(注41)名称はスペインのアラゴン地方で発見されたことに由来している。日本名「霰石(あられいし)」。主成分は炭酸カルシウム(CaCO3)、結晶系は斜方晶系、比重2.9の炭酸塩鉱物である。別名「なごみ石」とも呼ばれ、心身のバランスを整え、リラックスさせる効果があると伝承されている。
40億年前の冥王代の海で具体的にどのような構造が形成されていたのかは完全には明らかでないが、現在の環境との大きな違いが、少なくとも二つ存在し、それらが大きな影響を及ぼしていたことは確実である。
- 酸素が存在しなかった
- 大気と海洋のCO2濃度が現在よりもはるかに高かった
ことである。この二つの差異が、太古のアルカリ熱水孔をはるかに効果的な流通反応装置としたに違いなかった。
酸素がない環境下では、鉄は溶解性の高い第一鉄の形態で海に溶け込む。初期の海洋は溶解した鉄で満ち溢れていたことが明らかである。後にそれがすべて析出して、広大な『縞状鉄鉱層』となっているからである。この溶存する鉄の多くは、ブラックスモーカーの火山性熱水孔に由来する。
鉄がアルカリ熱水噴出孔で析出していたこともわかっている。この場合、鉄は水酸化鉄や硫化鉄として析出し、炭素やエネルギーの代謝を促す酵素に今も見つかるような触媒クラスター-フェレドキシン(注42)などのタンパク質-を形成していた可能性が高い。
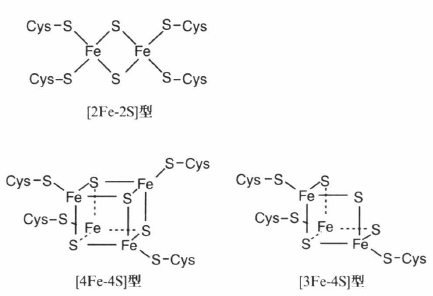
(注42)内部に鉄-硫黄クラスター (Fe-Sクラスター) を含む鉄硫黄タンパク質の一つであり、電子伝達体として機能する。ヘム(2価の鉄原子とポルフィリンから成る錯体である。通常、2価の鉄とプロトポルフィリンIXからなるプロトヘムであるフェロヘムのことをさすことが多い)を含まない非ヘムタンパク質(他にルブレドキシン、高電位鉄-硫黄タンパク質など)のひとつであり、動物から原核生物まで広く分布する。光合成、窒素固定、炭酸固定、水素分子の酸化還元など主要な代謝系に用いられる。なお、図中の「Cys」は「システイン(2-アミノ-3-スルファニルプロピオン酸)」を示すが、これはアミノ酸の1種である。
考察を進めると、
- 酸素が存在しなかったため、アルカリ熱水噴出孔の鉱物の壁は、触媒作用を持つ鉄鉱物を含んでいたことが推測され、さらに、反応性の高いニッケルやモリブデン(アルカリ流体に溶解していた)などの金属も加わっていた可能性がある。
- このような形で、アルカリ熱水噴出孔は真の化学反応装置に近づき、水素が豊富な流体が微細な通路を流れ、触媒作用を持つ壁が生成物を集め、濃縮・保持する一方で廃棄物を排出する構造を形成していた。
しかし、具体的にどのような反応が進行していたのかを考えると、ここで問題の核心に迫ることとなる。すなわち、
- 当時の高いCO2濃度が、反応の進行に大きな影響を与えていたのである。
現代のアルカリ熱水噴出孔は比較的炭素が乏しい。利用可能な無機炭素の大部分は、熱水孔の壁に「炭酸塩(アラゴナイト)」として析出しているからである。40億年前の冥王代には、CO2濃度は現在よりもはるかに高かったと考えられており、それが100倍から1000倍も高かった可能性もある。
原初の熱水孔で炭素がある程度多かっただけでなく、CO2濃度が高かったとすれば、海水の酸性度は高まり、炭素は炭酸カルシウムとして析出しにくくなっていただろう(現代の海洋ではCO2濃度の上昇により酸化が進み、サンゴ礁が脅かされている)。現代の海のpHはおおよそ8で、わずかにアルカリ性である。一方、冥王代の海はおそらく中性か弱酸性であった可能性があり、pH値は5~7であったかもしれない(注43)。
(注43)二酸化炭素は海面を通じて大気と海洋の間で活発に出入りする。海洋中に溶け込んだ二酸化炭素(CO2)は炭酸(H2CO3)となる(CO2+H2O ⇄ H2CO3)(式(1))。炭酸(H2CO3)は、海洋中では水素イオン(H+)が解離した炭酸水素イオン(HCO3–)や炭酸イオン(CO32-)との間で、(H2CO3 ⇄ H++HCO3–(式(2))、H++HCO3– ⇄ 2H++CO32-(式(3)))で表わされる反応により化学平衡の状態を保っている。大気中の二酸化炭素が増えると、海水に溶け込む二酸化炭素も増え、式(1)と式(2)の反応が右に進んで水素イオン(H+)が生成する。生成したH+の大部分は式(3)の反応が左に進むことにより消費されるが、一部のH+はそのまま残り、CO32-が減少する。結果としてH+が増加するためpHが低下し、海洋は酸性化することになる。
[補注:国土交通省気象庁、知識・解説、海洋酸性化の知識、海洋酸性化、https://www.data.jma.go.jp/gmd/kaiyou/db/mar_env/knowledge/oa/acidification.html]
以下の4つの要件が重要である。
- 高いCO2濃度
- 弱酸性の海
- アルカリ流体
- FeSをもつ熱水孔の薄い壁構造
これらの要素の組み合わせが、一般的には容易に発生しないような化学反応を促進する。
ところで、化学反応を制御する原理は大別して二つ存在する。「熱力学」と「反応速度論」である。熱力学は物質のどの状態が最も安定であり、時間に制約がなければどの分子が形成されるかを明らかにする。反応速度論は反応の速度に関連し、限られた時間内でどの生成物が形成されるかを決定する。
- 熱力学的に考えると、CO2は水素(H2)と反応してメタン(CH4)を生成する。
- これは熱を放出する発熱反応であり、一定の条件下では環境のエントロピーが増大し、反応が有利になる。適切な条件が揃えば、この反応は自然に起こるであろう。
- 必要な条件は、適度な温度と酸素の非存在などである。温度が高すぎると、CO2がメタンより安定になる。また、酸素が存在すると、酸素が水素と優先的に反応し、水が生成される。
40億年前、アルカリ性の熱水孔では適度な温度と酸素の存在しないことが、CO2とH2の反応を促進し、メタンが生成するのに有利な状況であったと考えられる。
今日でも、いくらか酸素があるものの、ロスト・シティーは少量ながらメタンを生成している。地球化学者のジャン・アメンドとトム・マッカラムはさらに踏み込んで、H2とCO2からの有機物の生成が、酸素のないかぎりアルカリ熱水噴出孔の条件では熱力学的に有利になる、と見積もった。そのような条件、すなわち、25~125℃では、H2とCO2からのあらゆる細胞バイオマス(アミノ酸、脂肪酸、炭水化物、ヌクレオチドなど)の生成は、実質的に『発エルゴン的』(エネルギーを発生するということ)と言える。したがって、そのような条件のもとで有機物はH2とCO2から自然に生成されるであろう。細胞の生成はエネルギーを放出し、全体のエントロピーを増すのである。
しかしながら、
- H2とCO2とは容易には反応しない。そこには反応速度論の障壁が存在する。
- つまり、熱力学的には自然に反応するはずであるが、何らかの別の障壁によって即座には起こらない状態になっているのである。
H2とCO2はそれぞれがほとんど活性を持たない。それらを無理やり反応させためるには、エネルギーの投入、反応を引き起こす何かが必要となる。その要素が加わると反応が始まり、まずは部分的に還元された化合物が生成される。CO2は電子をペアでしか受け取れない。電子が二つ加わると、ギ酸イオン(HCOO–)が生成される。さらに二つ加わるとホルムアルデヒド(CH2O)が生成され、さらに二つでメタノール(CH3OH)になり、最終的に二つ加われば完全に還元されてメタン(CH4)となる。もちろん生命はメタンだけで形成されているわけではなく、レドックス(酸化還元)状態ではホルムアルデヒドとメタノールの混合物に大体相当するような部分的に還元された炭素で構成されているのである。したがって、
CO2とH2から生命の誕生について考察する場合、反応速度論の二つの主要な障壁を認識する必要がある。
- 第一の障壁は、ホルムアルデヒドやメタノールに到達するためには必ず“克服しなければならない”ものである。
- 一方、第二の障壁は、それを“乗り越えてはならない”。H2とCO2を温かい環境へ誘導しても、反応が一気にメタンまで進行してしまえば、すべてがガスとして拡散し、そこで反応が終わってしまうからである。
- 細胞を生成するためには、第一の障壁を低下させ、第二の障壁を高める具体的な手法が必要となる。
この点が問題の核心である。「メタン生成菌」は、生育に必要なエネルギーと炭素をすべてH2とCO2の反応から得ているのに対し、我々はまだこの基本的な反応を効率的に推進することに成功していない。さらに困難を増しているのは、「どのような生物細胞も現われる前に、どのようにそれが形成されたのか?」という問題である。
しかし、可能な解決策の一つが考えられる。
プロトン・パワー
レドックス反応には、供与体(この場合はH2)から受容体(CO2)への電子の移動が関与している。そして、分子が自身の電子を運び出したがる傾向は、『還元電位』という概念により数値化される。すなわち、「還元電位」とは、
- 電子を奪われる傾向のある分子には、負の値が割り当てられる。
- 逆に、電子を求め、それをほぼどこからでも引き出す原子や分子には、正の値が割り当てられる。これは、負電荷を持つ電子を引きつける力と見なせば理解しやすい。酸素は電子を奪う傾向があり(電子を奪う相手を酸化する)、強い正の還元電位を持つ。
- これらの値はすべて、実際には『標準水素電極』という基準との相対値で与えられる。
要するに、
- 負の還元電位を持つ分子は自分の電子を剥ぎ取り、それに比べ正の還元電位をもつ分子に渡す傾向があり、その逆はないのである。
ここでH2とCO2に関する問題を考えてみよう。中性のpH(7.0)では、H2の還元電位は理論的にマイナス414mVである。H2が2個の電子を放出すれば、2個のプロトン(2H+)が残る。水素の還元電位はこの動的な均衡、すなわちH2が電子を失ってH+になる傾向と、2H+が電子を手に入れてH2になる傾向との均衡を反映している。この電子をCO2が取得した場合、それはギ酸イオンになるだろう。しかし、ギ酸イオンの還元電位はマイナス430mVである。それゆえ、電子をH+に渡してしまい、結果としてCO2とH2になる傾向が強い。ホルムアルデヒドの還元電位はさらに低く、およそマイナス580mVである。そのため、電子を持続的に保持しようとはせず、プロトンに渡してH2を生成する傾向が強い。したがって、
- pH7で考えれば、H2がCO2を還元できる可能性はない
のである。しかしもちろん、一部の細菌や古細菌はまさしくこの反応によって生きているので、可能性は必ず存在する。
では、どのような機構でこれが達成されているのだろうか。
分子の還元電位は、通常、pHすなわちプロトン濃度(水素イオン濃度)に依存する。その理由は単純で、電子が移動すると負電荷が移動することになるので、還元された分子(電子を受け取る分子)がプロトンも受け取ることが可能な場合、その生成物はより安定になる。それはプロトンの正電荷と電子の負電荷が釣り合うからである。
- 電荷を釣り合わせるために利用できるプロトンが多ければ多いほど、電子の移動はスムーズに進行する。結果として、還元電位も正にシフトする、すなわち電子のペアを受け取る傾向が強まる。実際、還元電位は、pHが1下がる(より酸性になる)ごとに約59mV増加する。
溶液の酸性度が高まるほど、電子がCO2に移動してギ酸イオンやホルムアルデヒドが生成しやすくなる。しかし残念ながら、これと全く同じ現象が水素でも起こる。溶液の酸性度が高まると、電子がプロトンに移動し、H2ガスが生成しやすくなる。したがって、
- 単にpHを変化させるだけでは何の効果もない。CO2をH2で還元することは不可能であるままである。
だがここで、「膜を挟んだプロトン勾配」を考えてみよう。膜の両側ではプロトン濃度-すなわち、酸性度-が異なるとする。この種の勾配は、アルカリ熱水孔でも観察される。
アルカリ熱流体は、細孔の迷路を通過しながらゆっくりと進行する。同様に、弱酸性の海水もそのように進行する。その中には、二つの流体が並行して流れる場所も存在し、
- CO2によって飽和した酸性の海水は、H2に富むアルカリ流体と、FeS鉱物という半導体を含む無機質の薄い壁によって隔てられている。
- H2の還元電位は、アルカリ性の条件では低下する。H2は電子を大いに奪われたがるため、そうして生成されたH+がアルカリ流体のなかでOH–と結合し、安定な水が形成される。
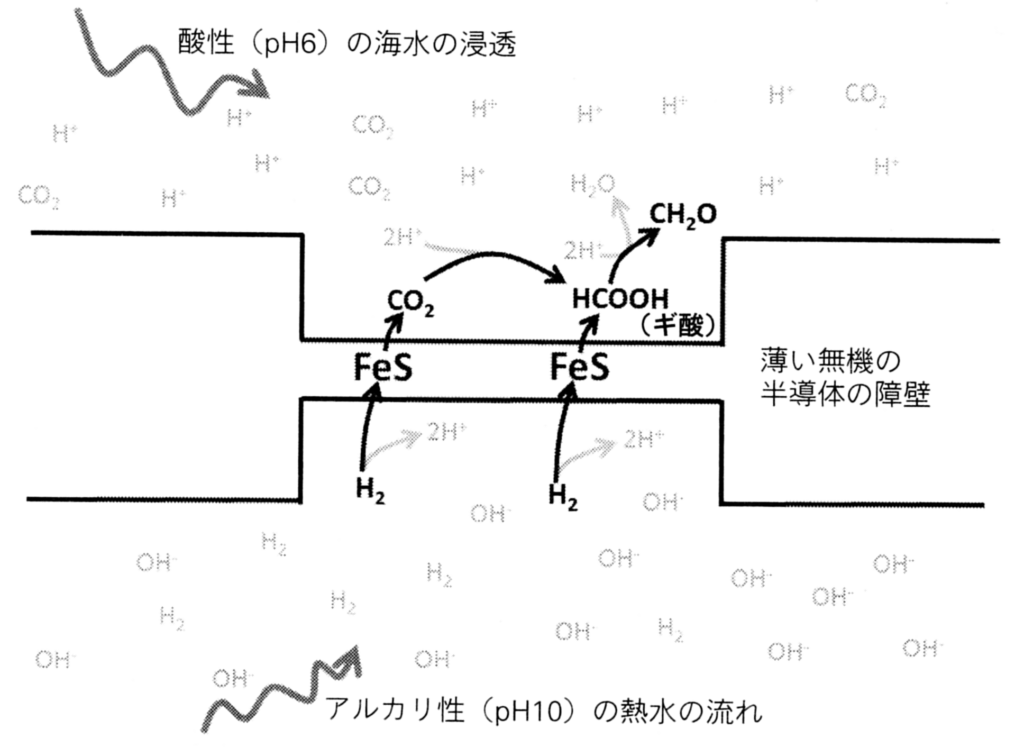
pH10では、H2の還元電位はマイナス584mVとなり、還元性が高い。一方、pH6で、ギ酸イオン(HCOO–)の還元電位はマイナス370mVであり、ホルムアルデヒド(CH2O)の還元電位はマイナス520mVである。すなわち、このようなpHの差が存在すれば、H2は比較的容易にCO2を還元してホルムアルデヒドを生成する。
ここで問題となるのは、電子がどのようにしてH2からCO2へ物理的に移動するのかという点である。
その答えは、細孔が無数に存在する熱水孔の構造にあり、薄い無機質の壁に含まれるFeS鉱物が電子を伝導する能力を持つ。その伝導能力は銅線には及ばないが、電子を移動させることは可能である。そのため、理論的には、アルカリ熱水孔の物理的構造はH2によるCO2の還元を促進し、有機物を生成するとされる(図50)。
この現象は検証可能であり、実際に、進化生化学者のニック・レーンらは、この反応を起こすための小さな卓上型反応装置を組み上げ、ギ酸イオンとホルムアルデヒドのほか、(リボースやデオキシリボースなどの)単純な有機物を合成したのである(注44)。
(注44)Barry Herschy, Alexandra Whicher, Eloi Camprubi, Cameron Watson, Lewis Dartnell, John Ward, Julian R. G. Evans, and Nick Lane, J. Mol. Evol. 79 (2014) 213–227.
どの種類の分子が形成され、それらがどのように形成されるのかについては後で議論することとし、とりあえずは、形成が可能であると仮定した場合、これらの有機物は、前述したように、『熱泳動』によって初めの濃度の数千倍に濃縮され、小袋や、もしかするとタンパク質などのポリマーも形成が促進されると考えられる。また、有機物が濃縮し重合するという予測も実験室で直接検証可能であり、ニック・レーンらの挑戦した実験では、初期の数段階は有望であると報告されている。ヌクレオチドと似た大きさの蛍光色素フルオレセインは、用意した流通反応装置で5000倍以上に濃縮され、キニーネはさらに濃縮されたという結果が出されている。
では、還元電位にまつわるこのこれらの事実は、何を示しているのだろうか。
「水素の還元電位がpHとともに落ちる」(すなわち、pHが上昇し、アルカリ性になると、還元電位がマイナスになり、その絶対値が大きくなる)という事実は、生命がこの宇宙で誕生するための条件について重要な意味を持つ。すなわち、
- アルカリ熱水の条件では、H2はCO2と反応し有機分子を生成すると予想されるが、他のほとんどの条件では、H2がCO2と反応し有機分子を生成することは起きないと考えられる、
ということである。
生命の起源となり得る条件として、事実上、他の全ての環境が排除される。
- 熱力学の原理に基づいて、細胞を一から作るには、制約のある流通反応システムの中で、反応性の高い炭素と化学エネルギーが、原始的な触媒の下で連続的に流れる必要があることが明らかとなった。
- そして、熱水噴出孔だけが、その必要条件を満たし、特に『アルカリ熱水噴出孔』は、必要な全ての条件を満たしている。
- アルカリ熱水孔のすばらしい特長は、アルカリ熱水には水素ガスが豊富にありながら水素はそのままではCO2と反応して有機分子を生成することがないという問題を抱えつつも、アルカリ熱水孔の物理的構造-薄い半導体の壁をはさんだ天然のプロトン勾配-が、有機物の生成を促し、さらに、そればかりでなく濃縮もする、という点なのである。
還元電位は生命が生まれるべき条件を狭めも広げもするが、これを検討する中で、生命の誕生を最も促進する条件が『アルカリ熱水孔』にあるという結論に至った。
「アルカリ熱水孔」の存在は可能性として高く評価され、その形成は水と鉱物『カンラン石』の化学反応に起因する。さらに、
- 「カンラン石」は宇宙において特に豊富に存在する鉱物であり、宇宙塵や、地球を含む惑星の形成素材である原始恒星系の円盤の大部分を占めている。
- カンラン石の『蛇紋岩化作用』(注37)は宇宙空間でも発生し、宇宙塵を水和(hydration、ある化学種へ水の分子が付加する現象)させる。我々の惑星が形成されるとき、この水は温度と圧力の上昇によって排出され、海を形成したとする説も存在する。
- カンラン石と水は、宇宙において特に豊富に存在する二つの物質である。
- また、CO2も同様に豊富で、太陽系の大半の惑星の大気や、他の恒星系の系外惑星の大気にさえ存在する。
- これら『岩石と水とCO2』は生命に必要な要素リストに挙げられ、湿潤な岩石惑星のほぼすべてに存在する。
- 化学と地質学の法則により、これらは、触媒となる細孔の薄い壁を挟んでプロトン勾配を持つ、温かいアルカリ熱水噴出孔を形成する。
- その化学反応が必ずしも生命の誕生を支援するわけではないかもしれないが、それは現在も、天の川銀河だけでも400億個も存在する地球型惑星で続行される実験である。
図表
図38 ユーリー-ミラーの実験
フリー百科事典『ウィキペディア(Wikipedia)』「ユーリー-ミラーの実験」(https://ja.wikipedia.org/wiki/%E3%83%A6%E3%83%BC%E3%83%AA%E3%83%BC-%E3%83%9F%E3%83%A9%E3%83%BC%E3%81%AE%E5%AE%9F%E9%A8%93)
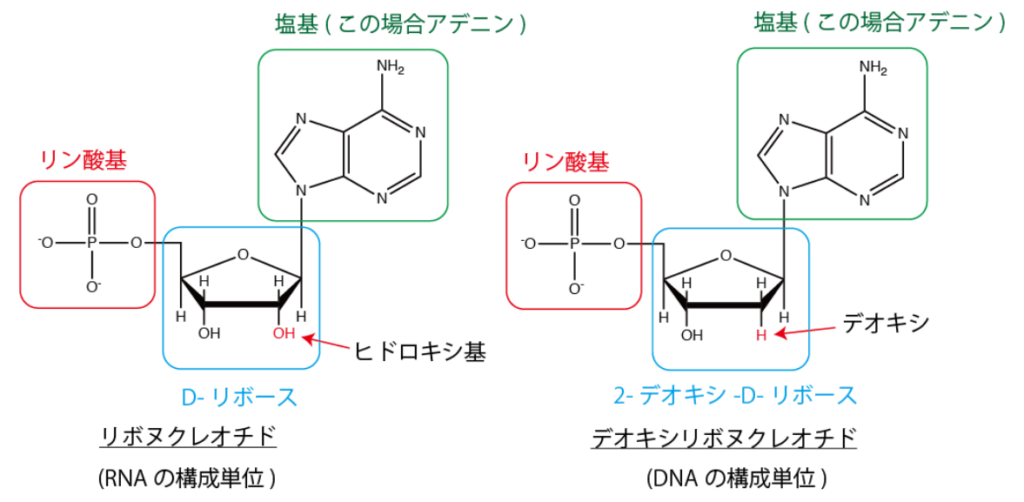
図39 ヌクレオチドの構造
生命系のための理工学基礎、生化学、ヌクレオチドと核酸・DNAとRNAの構造を解説
(https://rikei-jouhou.com/polynucleotide/)
図40 ATPの加水分解反応
長近未来メディア教育研究所 谷川建治「生物における時間の意義を考える」
(https://sites.google.com/site/lifetimearrow/home?authuser=0)
3)動力装置と燃料の話・基礎編 図1ATP加水分解反応の模式図
(https://sites.google.com/site/lifetimearrow/senmou-to-nenryou?tmpl=%2Fsystem%2Fapp%2Ftemplates%2Fprint%2F&showPrintDialog=1)
図41 アセチルリン酸の化学構造
健康用語WEB事典・アセチルリン酸(acetylphosphate)
(https://health.joyplot.com/HealthWordsWiki/?%E3%82%A2%E3%82%BB%E3%83%81%E3%83%AB%E3%83%AA%E3%83%B3%E9%85%B8)
図42 コロイド中のリン脂質のとる形態
フリー百科事典『ウィキペディア(Wikipedia)』「脂質二重層」
(https://ja.wikipedia.org/wiki/%E8%84%82%E8%B3%AA%E4%BA%8C%E9%87%8D%E5%B1%A4)
図43 RNAの化学構造
一般社団法人日本生物物理学会、生物物理について・核酸構造
(https://www.biophys.jp/highschool/B-03.html)
図44 鉄硫黄クラスター
Journal of Geography(Chigaku Zasshi)129(6)853870,2020(https://www.jstage.jst.go.jp/article/jgeography/129/6/129_129.853/_pdf)
図45 海洋の発散型境界(中央海嶺の例)
大鹿村中央構造線博物館・学習資料・マグマのでき方1:中央海嶺とホットスポットのマグマ
(https://mtl-muse.com/study/kashio-spring/magma1/)
図46 蛇紋岩
HAGS (ハグス)・リノベーション用語集・「蛇紋岩(じゃもんがん)」とは何か?
(https://hags-ec.com/column/503-special-glossary/)
図47 地球深部でのカンラン石の変化
国立研究開発法人海洋研究開発機構、プレスリリース・話題の研究 謎解き解説・理論予測されて
いたカンラン石組成の新物質を隕石から発見
(https://www.jamstec.go.jp/j/about/press_release/quest/epsilon/index.html)
図48 ロスト・シティー
Alexander S. Bradley“Expanding the Limits of Life”,SCIENTIFIC AMERICAN, December
2009
図49 フェレドキシンの酸化還元中心の3型
光合成事典・フェレドキシン
(https://photosyn.jp/pwiki/index.php?%E3%83%95%E3%82%A7%E3%83%AC%E3%83%89%E3%82%AD%E3%82%B7%E3%83%B3#:~:text=%E3%83%95%E3%82%A7%E3%83%AC%E3%83%89%E3%82%AD%E3%82%B7%E3%83%B3%5Bferredoxin%5D%20%E2%80%A0,%E7%A1%AB%E9%BB%84%E3%82%AF%E3%83%A9%E3%82%B9%E3%82%BF%E3%83%BC%E3%81%8C%E5%AD%98%E5%9C%A8%E3%81%99%E3%82%8B%EF%BC%8E)
図50 アルカリ熱水孔での有機物生成のモデル
「生命、エネルギー、進化THE VITAL QUESTION Why Is Life the Way It Is?」
ニック・レーン Nick Lane著、斉藤隆央訳、株式会社 みすず書房、2016年9月13日 第1刷、
2018年3月9日 第10刷、図14「H2とCO2からの有機物のできかた.B」、135頁
