
第三部 生命の誕生
はじめに
約40億年前に大規模な原始海洋が形成され、ついに「生命の誕生」の舞台が設えられた。
“無から有”への転換、不連続的な発現である『宇宙の誕生』に劣らず、『生命の誕生』もそれに比肩しうるほどの劇的な事象である。
前の部で『太陽系の誕生』と『地球の誕生』という事象を眺めた。これらも実際は、それまでにはなかったものを生み出すという意味では、革新的な事象ではある。しかし、その誕生の物語は“無から有”というレベルの不連続的な発現ではなく、ある特定な環境下で連続的な現象の積分的な積み重ねから実現されたものである。
生命の誕生も、実は、不連続的な発現ではなく、ある特定な環境下で連続的な現象の積分的な積み重ねから実現されたものであることが解明されつつある。そればかりでなく、最近の優れた研究により明らかにされつつあることは、
- 生命が『エネルギーの流れ』に深く関係しているということである。すなわち、「エネルギーは生命の進化の要」であり、エネルギーを方程式に持ち込んで初めて生命進化の特質が理解できる。しかも、進化ばかりではなく、このエネルギーと生命の関係は生命の始まりにまで遡るのである。
- 『生命』の根本的な特質、それは、たゆまず活動する惑星に生じた不均衡から必然的に現われたものなのである。生命がエネルギーの流れによって誕生し、『プロトン勾配』が細胞の出現の鍵を握っていたのである。
以下に、そのことを具体的に説明してみよう。
第1章 生命
生命とはなにか?
2020年にパンデミックとなった新型コロナウイルス感染症は大流行した。インフルエンザは冬になると新型コロナウイルス感染症に追随して現れる。ノロウイルス、風疹ウイルス、肝炎ウイルスなどの『ウイルス』は各種の感染症を引き起こす厄介な存在である。この厄介なウイルスは生物なのだろうか。
実は、ウイルスは生物ではない。なぜなら、それ自身の『代謝』が働いていないからである。宿主の力に全面的に依存している(注15)。
(注15)現在、多くの生物学者が認めている生物の定義とは次の3つの条件、①外界と膜で仕切られている、②代謝(物質やエネルギーの流れ)を行う、③自分の複製を作る、を満たすものである。これは意外とシンプルな定義であり、これだけで生物を定義できることが不思議に思えるかもしれない。しかし、現在のところ、この3つの条件をすべて満たすものは生物だけである。
ここで『代謝(metabolism)』という重要な概念が出てきたが、「代謝」とは、生命の維持のために有機体が行う、外界から取り入れた無機物や有機化合物を素材として行う一連の合成や化学反応のことであり、『新陳代謝』の略称である。これらの経路によって有機体はその成長と生殖を可能にし、その体系を維持している。
「代謝」は大きく『異化』 と『同化』の2つに区分される。
- 「異化」は物質を分解することによってエネルギーを得る過程、例えば「細胞呼吸」がこれに該当する、
- 「同化」はエネルギーを利用して物質を合成する過程、例えば「タンパク質・核酸・多糖・脂質の合成」がこれに該当する。
しかしながら、真に、「代謝」の活動は生命にとって不可欠な特性なのだろうか。生命を定義し、生物と無生物を明確に区別する要素なのだろうか。
環境からの助けを必要とするのか?
ウイルスは周囲の環境を活用して自己の複製を作り出す。これは私たちが行っていることと酷似している。他の動物や植物を摂取し、酸素を吸い込む私たちも、ビニール袋で頭部を覆い、環境から分離すると、僅かな時間で命を絶つ。私たちもウイルスと同様に、環境に対して寄生的な存在であると言えるだろう。同様に植物も、私たちが植物を必要とする程度とほぼ同等に、私たちを必要としている。光合成により自己の有機物を生成し、生育するために、植物は日光、水、二酸化炭素(CO2)を必要とする。乾燥した砂漠や暗い洞窟では成長できず、CO2が不足していても成長は困難である。植物に必要なこのガスが不足しない理由は、動物、菌類、さまざまな細菌が持続的に有機物を分解し、消化し、燃焼させ、最終的に大気中にCO2として排出しているからである。我々が化石燃料を全て燃やしてしまうことが地球に悪影響を及ぼす可能性がある一方で、植物にとってはCO2が豊富であれば成長が活発になるため、この事象を感謝すべき事情と捉えることも可能である。結果として、植物も私たちと同様に、環境に寄生していると解釈できる。
この視点で見れば、植物と動物とウイルスの差異は、それぞれの環境から受け取るものの差に過ぎないことになる。
細胞内では、ウイルスは全てを満たすことのできる「完璧な子宮」の中で大切に育てられている。彼らはほとんどのものをそぎ落とした姿でも生きていけるが、それは身のまわりの環境が非常に豊かなためにほかならない。一方で、植物は身のまわりの環境にきわめて低い要求しかしない。光、水、空気があればほぼどこでも成長することができる。外部要因が非常に少なくても生存するため、内部機構は必然的に複雑化している。自己の生化学的メカニズムによって、植物は成長に必要なものを全て、空気中から合成することが可能である。私たち人間は、このようなウイルスと植物の間の存在と言えるだろう。食べるという普遍的な要求を超えて、私たちは食事に特定のビタミンを必要としており、それがないと壊血病などの恐ろしい病気になる。ビタミンは私たちが単純な前駆体から自ら合成できない化合物であり、祖先が持っていたそれを合成する生化学的メカニズムを我々は既に失っている。このビタミンによる外部からの援助が無いと、私たちは宿主を持たないウイルスと同じ運命をたどることになるだろう。
このように私たちが皆環境からの助けを必要とするのであれば、残る疑問は「どれだけ必要なのか?」ということだけになる。
環境への依存度
ウイルスや『レトロトランスポゾン』(注16)など、DNAの一部に寄生する『パラサイト(寄生体)』は、安全な宿主から離れず、全ゲノムに自己を複製する。レトロトランスポゾンもウイルスも、いずれも身のまわりの生物学的環境を利用して、みずからのコピーを作る能力(自己複製能力)を有する。言い換えれば、無生物と生物は連続しており、その間で線引きを試みることは意味をなさない。生命の定義の大半は生体そのものに目を向けていて、環境への寄生については往々にして無視している。たとえばNASAによる生命の「暫定的な定義」では、生命とは「ダーウィン進化可能な自立した化学的システム」とされている。この定義にウイルスは含まれないかもしれないが、「自立した」という曖昧な言葉の解釈にもよるだろう。
(注16)ゲノムの中を動くことのできる遺伝子のことを『トランスポゾン(可動遺伝因子)』という。トランスポゾンにはその動き方によって、「DNA型トランスポゾン」と「レトロトランスポゾン(RNA型トランスポゾン)」に分けられる。「レトロトランスポゾン」は、供与DNAが転写されてRNA(一次コピー)となり、RNAがさらにコピーされ、DNAが合成される(二次コピー)。コピーされたDNAが挿入されるので、レトロトランスポゾンはコピー&ペーストであるのが特徴で、供与DNAにあったレトロトランスポゾンは動かず、元の場所に残る。一方、「DNA型トランスポゾン」は、DNAのまま動けるのでカット&ペーストなのが特徴である。
なお、「トランスポゾン」がゲノムの中を移動する仕方については、例えば以下のサイトに模式的に分かりやすく示されている。https://researchmap.jp/blogs/blog_entries/view/76287/67b931a8e4081b49fc27e3b9398df10d?frame_id=527881
いずれにせよ、生命が環境に依存することが重視されているわけではない。環境は、まさにその本質から言って、生命の外のものに見えるが、決してそうではなく、両者はいつでも密接に結びついているのである。
生命が適応している環境から分離されたらどうなるだろうか。私たち人間はおそらく生存できず、死を迎えるだろう。私たちには生と死の二択しか存在しないように思えるが、それが常に当てはまるとは限らない。宿主細胞の資源から切り離されても、ウイルスはすぐに崩壊して「死ぬ」ことはない。世界からあれこれ奪われてもかなり耐えられるのだ。たとえば、1ミリリットルの海水中には細菌の10倍のウイルスが存在し、その時を待っている。ウイルスが崩壊に耐える能力は、仮死状態で保たれ長い年月を過ごすことができる細菌の『芽胞』を思い起こさせる。芽胞は代謝活動を一切行わず、何千年もの永久凍土やさらには宇宙空間で生き延びる能力を持っている。また、芽胞だけでなく、種子やクマムシのような生物も、完全な乾燥状態や、ヒトを殺す量の1000倍の放射線、海底の途方もない圧力、また宇宙の真空といった極限環境に耐えることができる。
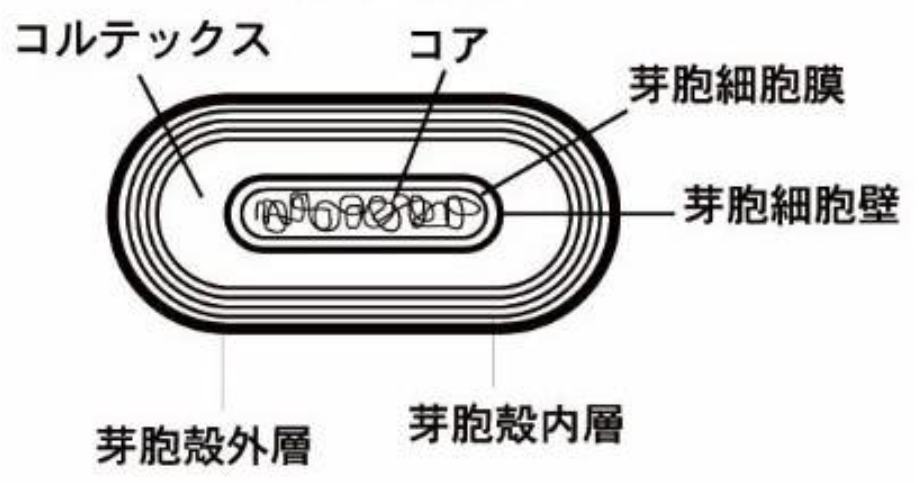
ウイルスや芽胞やクマムシが、熱力学第二法則により崩壊すべきとされるにも関わらず、なぜ崩壊しないのか。最終的には崩壊するかもしれないが、それがない場合、彼らは生命活動を行わない状態であるにも関わらず、驚くほど安定している。
これは、『生命(Life)』と『生(Living)』の違いについて重要なことを教えてくれている。
- 芽胞は、多くの生物学者によって生物に分類されるものの、厳密には生きていない状態である。それでも、蘇生する能力を持ち、再び生命活動を行うことができるため、死んでいるわけではない。
- ウイルスも、適切な環境が整えば再び自己複製する能力を持つ。クマムシも同様である。
- 『生命』は、遺伝子や進化によって決定される構造であるが、「生」は成長や繁殖といったプロセスに加えて環境にも依存しており、これは構造と環境の相互関係に他ならない。
エネルギー、エントロピー、構造
『熱力学第二法則』によれば、『エントロピー(無秩序さ)』は必ず増大するのだから、一見したところ芽胞やウイルスがそのように安定しているというのは不思議に思える。
生命とは違ってエントロピーは具体的に定義されており、測ることができ、単位はジュール/ケルビン/モルである。そこで、芽胞を粉々に砕いてエントロピーの変化を調べてみよう。すりつぶして構成要素の分子にまでばらばらにし、エントロピーの変化を測ってみると、エントロピーは増えているに違いない、と思われる。というのは、しかるべき条件になれば再び成長できる見事に秩序立っていた系が、ランダムで機能しないかけらの集まりになったのだから、当然、エントロピーは高いと考えられる。
しかし、生体エネルギー学者テッド・バットリーの精密な測定結果によれば、エントロピーはほとんど変化していなかった。なぜなら、エントロピーには、芽胞以外のものも関与するからである。環境も考慮しなければならず、それにもある程度の無秩序さがあるのである。
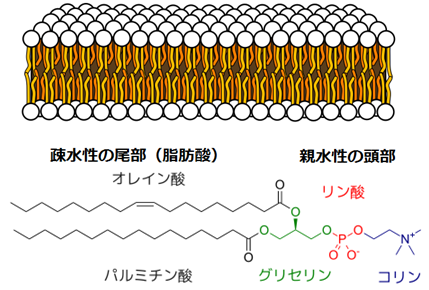
芽胞は、相互作用するパーツがきっちり収まってできている。その中では、油性(脂質)の膜が、分子間に働く物理的な力によって自然に水を遠ざける。油性の脂質を水に混ぜて振ると、おのずから薄い『二重層』になり、これが水性の小胞を覆う生体膜となる。それが最も安定した状態だからである。すなわち、物理的な引力と斥力から、油と水は、相互にではなく自分自身と作用したがるのである。タンパク質もほぼ同じような振る舞いをする。電荷を多く持つものは水に溶け、電荷のないものは油とよく作用する『疎水性』を示す。つまり、水を嫌うのである。油性の分子が集まって落ち着き、電荷を持つタンパク質が水に溶けると、エネルギーが放出される。それが物質の、物理的に安定した、低エネルギーの、「心地よい」状態なのだ。エネルギーは熱として放出される。
熱は分子の運動で、押し合いへし合いする分子の無秩序さであり、結果的にエントロピーをもたらすものなのである。したがって、油と水が分離して熱が放出されると、エントロピーが増大する。すると、全体のエントロピーの観点からは、こうした物理的相互作用をすべて考慮すれば、秩序立った膜が細胞を取り囲むのは、混和しない分子がランダムに混じった状態よりも、エントロピーが高いということになる。
物理的に心地よい状態が環境にエネルギーを熱として放出するのなら、物理的に心地よくない状態は逆のこと、すなわちエネルギーを環境から吸収し、環境のエントロピーを下げて環境を冷やすことになるのである。芽胞の場合にこうしたすべてを考慮にいれると、全体のエントロピーはほとんど変化しない。
分子のレベルでは、ポリマーの構造がエネルギーを局所的に最小化し、余剰のエネルギーを熱として環境に放出して、環境のエントロピーを増大させる。タンパク質はできるだけ低いエネルギーをもつ形状に自然に折りたたまれる。その疎水性の部分は、表面の水から離れた場所に埋まっている。電荷は引きつけ合ったり斥け合ったりするので、正電荷は負電荷との釣り合いで決まった場所にとどまり、タンパク質の立体構造を安定させている。こうしてタンパク質は、必ずしも有用ではなくても、特定の形状に自然に折りたたまれる。
なにより驚くのは、タンパク質の構成要素であるばらばらのアミノ酸からなる無秩序なスープと、見事に折りたたまれたタンパク質とで、全体のエントロピーにほとんど差がないことかもしれない。たたまれたタンパク質を広げると、アミノ酸のスープにより近い状態に戻り、エントロピーが増す。しかしそうすると、疎水性のアミノ酸を水にさらすことにもなり、この物理的に心地よくない状態は外部からエネルギーを吸収し、環境のエントロピーを減らして周囲を冷やす。
生命がエントロピーの低い状態、つまり無秩序なスープよりも組織化されているという考えは、厳密には正しくない。生命の秩序と組織を、環境の無秩序さの増大が上回っているからである。
それでは、エルヴィン・シュレーディンガーが「生命は環境から負のエントロピーを吸い上げる」と述べた(注17)が、それはどういう意味なのだろうか。
- 彼はその言葉で、「生命は周囲からどうにかして秩序を引き出す」のだと述べたかったのである。
(注17)エルヴィン・シュレディンガー著「生命とは何か」Erwin Schrodinger “What is life?”
[訳]岡小天・鎮目恭夫、岩波新書 1951・1975
実は、たとえアミノ酸のスープがきっちり折りたたまれたタンパク質と同等のエントロピーであっても、ふたつの意味で、タンパク質のほうが生じにくく、それゆえエネルギーを要するのである。第一に、アミノ酸のスープは自然に結合して鎖を形成することはない。タンパク質はアミノ酸が連なった鎖だが、アミノ酸は本来反応性が低い。アミノ酸を結合させるには、生体細胞はまずそれを活性化しなければならない。そうして初めて、反応して鎖ができるのである。このとき、最初の活性化で使われたのとほぼ同じ量のエネルギーが放出される。タンパク質が自分を折りたたむときに放出するエネルギーは熱として失われ、環境のエントロピーを増大させる。だから、
同等に安定なふたつの状態のあいだにも『エネルギー障壁』(活性化エネルギー)がある。エネルギー障壁から、タンパク質の形成が難しいと言えるのと同じく、タンパク質の分解にも障壁がある。タンパク質を壊して構成要素に戻すのには、ある程度の労力(と消化酵素)が要る。
反応性の高い構成要素が十分にあれば、そうした大きな構造のほうがとくに安定した状態となる。そして実際の問題は、「その反応性の高い構成要素はどこから生まれてくるのか?」ということに行き着く。
ここから第二の問題も導かれる。活性化したアミノ酸はおろか、アミノ酸のスープさえ、今日の環境でも存在が確かだとは言えない。そのまま放置すると、やがて酸素と反応してもっと単純なガスの混合物-二酸化炭素、窒素酸化物、水蒸気-に戻ってしまうだろう。つまり、そもそもアミノ酸ができるのにエネルギーが要り、そのエネルギーはアミノ酸が分解されるときに放出されるのだ。だからわれわれは、飢えてもしばらくは、筋肉のタンパク質を分解し、それを燃料として生き延びることができる。このエネルギーは、タンパク質そのものではなくそれを構成するアミノ酸の燃焼によって得られるものだ。したがって、種子も芽胞もウイルスも、今日の酸素の豊富な環境では完全に安定してはいない。それらの構成要素はゆっくり時間をかけて酸素と反応し(酸化され)、ついには構造や機能が蝕まれて、適切な条件でも蘇生できなくなる。種子は死ぬのだ。しかし取り巻く気体を変え、酸素を寄せつけなければ、いつまでも安定する。生物は酸素に富む地球環境によって平衡状態でなくなっているので、積極的に進行を防がないかぎり酸化されやすいのである。このように、
通常の環境(酸素の存在下)では、二酸化炭素や水素などの単純な分子から、アミノ酸のほか、ヌクレオチドなど生物の構成要素を作るのに、エネルギーが要る。また、それらをつなげて長い鎖、タンパク質やDNAなどのポリマー、にするのにもエネルギーが要るが、エントロピーの変化はほとんどない。それが『生』というものだ。
- 新たな要素を作り、それをすべてつなげ、成長し、殖える。成長とは、細胞の内外に積極的に物質を輸送することでもある。こうしたことを行うには、エネルギーの継続的な流れが必要になる。
- シュレーディンガーはそれを『自由エネルギー』と呼んだ。彼が念頭に置いた方程式は象徴的なもので、エントロピーと熱を自由エネルギーに関係づけている。
ΔG=ΔH-TΔS
ギリシア文字のΔ(デルタ)は、変化を示しているが、ΔGは『ギブスの自由エネルギー』の変化で、19世紀の偉大な孤高のアメリカ人物理学者J・ウィラード・ギブスの名にちなんでいる。このエネルギーによって、筋肉の収縮などの機械的な仕事や細胞内の現象が「自由」に引き起こせる。
ΔHは熱の変化で、環境に放出され、その環境を暖めるので、環境のエントロピーを増大させる。熱を環境に放出する反応は、系そのものを冷やす。反応前よりも系内のエネルギーは減るからだ。すると、系から環境に熱が放出される場合、系のΔHは負の符合を持つ。
Tは温度で、その重要性はあくまで環境に応じて決まる。ある量の熱を冷たい環境に放出すると、まったく同じ量の熱を暖かい環境に放出するよりも、その環境に大きな影響を及ぼす。相対的なインプットが大きくなるからである。
最後にΔSは、系のエントロピーの変化である。系のエントロピーが減ると、これは負の符合をもつようになって秩序を高め、逆にエントロピーが増すと正になり、系は無秩序さを増す。
全体として、どんな反応であれ自然に起こるためには、自由エネルギーΔGは負でなければならない。これは『生』を構成するすべての反応の総和についてもやはり言える。つまり、ΔGが負の場合にのみ、反応が自発的に起こるのである。そうなるためには、
- 系のエントロピーが増大する(系の無秩序さが増す)か、
- 系からエネルギーが熱として失われるか、
- あるいは両方が起きるか、
の必要がある。
それゆえ、局所的なエントロピーは、ΔHがそれ以上に負である(つまり大量の熱が環境に放出される)かぎり、減少しうる-系は秩序を増しうる-のだとも言える。結局、
- 成長と繁殖、すなわち『生』、を促すには、なんらかの反応が環境に絶えず熱を放出し、環境の無秩序さを増大させなければならないのである。
満天に輝く星々は、莫大な量のエネルギーを宇宙に放出することで、秩序立った存在の代価を払っている。私たち自身の場合、呼吸という不断の反応による熱を放出することで、持続的な存在の代価を払っている。われわれは絶えず酸素のなかで食物を燃やし、環境に熱を放出している。その熱損失は無駄ではない。生命が存続するためには絶対に必要なのである。
生体細胞で起こる現象はすべて自発的で、しかるべき起点を与えてやればひとりでに進行する。ΔGはつねに負だ。エネルギーはずっと下降線を辿る。しかしこれは、起点が非常に高いところになければならないことを意味する。
タンパク質を作るとしたら、起点は、活性化したアミノ酸を小さなスペースにたっぷり詰め込んだ、とんでもない集合体となる。そうしたアミノ酸は、結合し折りたたまれてタンパク質を形成するときにエネルギーを放出し、環境のエントロピーを増大させるだろう。
活性化したアミノ酸は、適度に反応性の高い前駆体が十分にあれば、自発的にできる。また、そんな適度に反応性の高い前駆体も、反応性の高い環境が与えられれば、自発的に形成される。だから結局のところ、
- 成長のパワーは環境の反応性に由来するもので、私たちの場合は食物と空気という形で、植物の場合は光子という形で生体細胞を絶えず流れている。
- 生体細胞はこの継続的なエネルギーの流れを成長につなげ、分解されてしまう傾向に打ち勝つ。
それは、遺伝子がある程度指定する精巧な構造によってなされている。だが、そのような構造が何であろうと、それ自体が、成長と複製、自然選択と進化のもたらす結果であり、
- そうした原因のどれもが、環境のどこかから絶えず流れ込むエネルギーなくして起こりえないのである。
- 生物が生きるのには、途方もない量のエネルギーが必要となる。
あらゆる生体細胞が用いるエネルギーの媒体『エネルギー通貨』は、『ATP(adenosine triphosphateアデノシン三リン酸)』という分子である。ATPは、特定のタンパク質が安定した状態から別の状態へ変化するためのエネルギーを供給する。つまり、タンパク質のある安定した立体構造から別の立体構造に変化させるのである。しかし、この変化後、タンパク質は再び動きを止める。構造を元に戻すためには、再度ATPが必要となる。
細胞内には多数のタンパク質が存在し、それらはATPによって動かされる。それゆえ、一つの細胞は毎秒約1000万個のATP分子を消費していると言える。その数は驚くほど多い。人体には約40兆個の細胞が存在し、それらの細胞が消費し再生成するATPの総量は1日あたり約60〜100キログラムとなり、これは我々自身の体重とほぼ同等の量である。しかし、実際には、我々の体内に存在するATPは約60グラムであり、この事実から、各ATP分子が1分に一度か二度補充されていることが示唆される。
その補充のメカニズムは以下のとおりである。
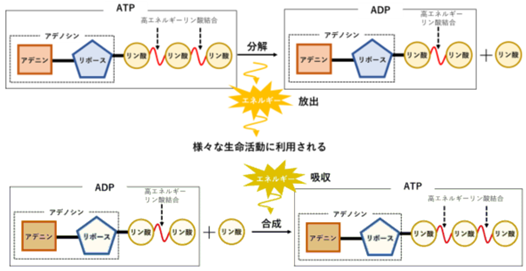
- ATPが分解される際、立体構造の変化を引き起こす自由エネルギーを放出する。同時に、ΔGを負に保てるだけの熱も放出する。ATPは通常、『ADP(adenosine diphosphateアデノシン二リン酸)』と『無機リン酸(PO43-)』の二つの部分に分割される。この無機リン酸は、肥料に使われているものと同じで、ふつうはPiと表示される。
- そして、ADPとPiから再度ATPを形成するのにもエネルギーが必要となる。呼吸によって得られるエネルギー(食物と酸素の反応によって放出されるエネルギー)が、ADPとPiからATPを再生するために用いられる。
- これがすべてだ。このエネルギーの循環こそが、生命のエネルギー代謝サイクルの全体像である。
ADP+Pi+エネルギー ⇋ ATP
私たちヒトのATP消費量は比較的少ない。例えば、20分ごとに分裂する「大腸菌」のような細菌は、成長のためのエネルギーとして、1回の細胞分裂で約500億個のATPを消費する。これは細胞1個の重さのおよそ50~100倍にもなり、我々のATP合成速度のほぼ4倍に相当する。このエネルギー消費量をワットで換算すると、生命のエネルギー消費量は、以下のとおり信じがたいものとなることがわかる。
- 我々ヒトは、体重1グラムあたり約2ミリワットのエネルギーを消費している。したがって、体重65キログラムの平均的な人間は、約130ワットのエネルギーを消費しており、これは100ワットの標準的な電球を少し上回る程度である。
- この数値は大きく感じられないかもしれないが、1グラムあたりでは太陽が消費するエネルギーの1万倍にもなる。
- 実は、どの瞬間においても、太陽はほんの一部しか核融合を起こしていないということになる。
- 生命のエネルギーの使い方は、ローソクというよりも、発射台のロケットに似ている。
したがって、理論的に見れば、生命は謎めいたものではなく、自然法則を破っているわけでもない。生体細胞が毎秒消費するエネルギーの量は非常に大きいが、地球に降り注ぐ太陽エネルギーの量はそれを大幅に上回る。太陽のエネルギーは1グラム当たりのパワーが少ないが、その大きさがそれを補っている。このエネルギーの一部が生化学的メカニズムを駆動するのであれば、生命はほぼあらゆる方法で活動することが可能であると考えられる。エネルギーの使い方には、根本的な制約が存在しないと思われる。
しかし、驚きを禁じ得ない事実として、地球上の生命はエネルギー利用の観点から見ると、極めて大きな制約を受けているのである。
具体的には、生命のエネルギー利用に関して二つの意外な特徴が存在する。
- まず一つ目は、あらゆる細胞のエネルギーが『レドックス反応』と称される一種類の化学反応から得られているという事実である。この反応は、「分子間で電子を受け渡す現象」である。
- 『レドックス(Redox)』とは、「酸化還元(reduction and oxidation)」を指す語であり、これは一つまたは複数の電子が供与体から受容体へ移る現象を表す。電子を供与した体は酸化されると考えられる。

この現象は、鉄などが酸素と反応する際に起こり、鉄は酸素に電子を供与し自身が酸化される。電子を受け取った物質-この場合は酸素-は還元される。例えば、呼吸や燃焼において、酸素(O2)が還元されて水(H2O)になる。酸素原子が電子2個とプロトン(陽子)2個を取得し、電荷のバランスを保つからである。
この反応はエネルギーを熱として放出し、反応を起こす系そのもののエネルギーは減少する。特に鉄や食物と酸素との反応では、大量のエネルギーが燃焼のように放出される。
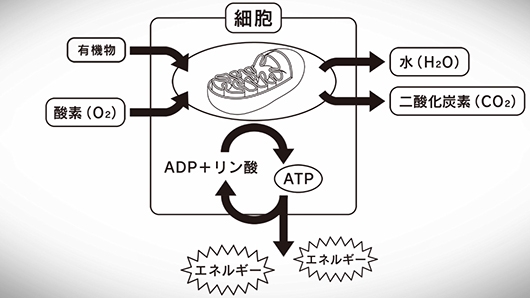
呼吸は、この反応で放出されるエネルギーの一部を、少なくともATPが再び分解されるまでの短い期間、ATPの形で保存する。分解時に、ATPのADP-Pi結合に残ったエネルギーは熱として放出される。結局のところ、呼吸と燃焼は等価であり、その過程のわずかな遅延が『生命』として認識される存在である。
一連のレドックス(酸化還元)反応は、結局のところ運び手の連鎖を辿る電子の移動であり、導線を流れる電流にも似ている。呼吸ではそのような電子の移動を行っているのである。
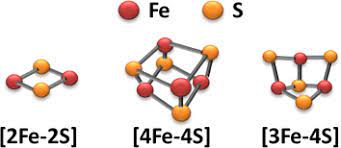
食物から奪われた電子は、酸素に直接渡されず、「飛び石」(通常は呼吸系タンパク質に含まれるイオン化した鉄原子(Fe3+)の一部であり、しばしば『鉄硫黄クラスター』という小さな無機結晶に含まれる)に移る。そこから電子が、とてもよく似ているがわずかに電子の「必要性」が高いクラスターへ跳ぶ。電子が次のクラスターに引き寄せされて渡るとき、クラスターはまず還元され(電子を受け取るのでFe3+がFe2+になる)、それから酸化される(電子を失うのでFe3+に戻る)。このプロセスを約15回以上繰り返すと、電子は最終的に酸素に到達する。
植物の光合成と動物の呼吸のように、一見したところ共通点がほとんどないように思える成長の手段も、そのような『呼吸鎖』を辿る電子の移動が関わっているという点で、実は基本的に同じなのである。
生命は熱エネルギーや力学的エネルギー、放射能、放電、紫外線放射など、想像しうる種類のエネルギーを用いて機能する可能性がある。しかしながら、驚くべきことに、すべての生命体はほとんど同様の呼吸鎖を介した「レドックス反応」でエネルギーを獲得している。
なぜなのだろうか。
生命のエネルギーに関する第二の意外な点は、ATPの結合にエネルギーを保存する具体的なメカニズムである。
- 生命は単純な化学反応を使うのでなく、薄い膜を隔てた『プロトン勾配』(注18)という手段によってATPの生産を促進する。
- 今日では、『プロトン勾配』が生じる分子レベルのメカニズムが詳細に解明されており、また、プロトン勾配が、地球上の生命に普遍的に見られることもわかっている。「プロトンのパワー」は、普遍的な遺伝コードであるDNAと同じくらい、生命に不可欠の要素なのである。
(注18)「プロトン勾配」とは、「電気化学ポテンシャルの勾配であり、膜を越えて移動する「プロトン(水素イオン)」についてのものである。勾配は、「化学的勾配」(膜を挟んだ溶質濃度の差)と「電気的勾配」(膜を挟んだ電荷の差)という2つの部分から構成される。透過性の膜を挟んだ両側のプロトン濃度が不均等であるときには、プロトンは高濃度側から低濃度側へ単純拡散によって膜を越えて移動する。プロトンは電荷を持っているため、膜を挟んで電位も形成される。膜を挟んで電荷が不均等に分布している場合、膜の両側で電荷が均等となるまでプロトンの拡散を駆動する力が電位差によって生み出される。
ところが、生命体のエネルギー生成を担うこのメカニズムがどのように進化を遂げたのかについては、まだほとんど理解されていない。
理由はどうあれ、地球上の生命は、エネルギー生成のメカニズムとしてありうるもののうち、驚くほど限られた特異な一部 -『レドックス反応』と『プロトン勾配』- しか利用していないように見える。これは歴史的な偶然を示しているのか、それともこれらの方法が他のどの方法よりも優れていたために広く普及したのか、最終的な疑問として、これらが唯一の可能な方法なのか―これらの疑問はまだ解答を待っている。
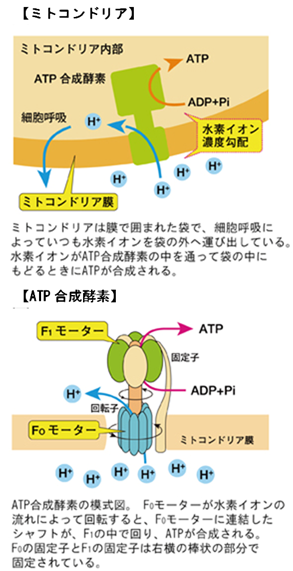
ミトコンドリア内のATP合成酵素
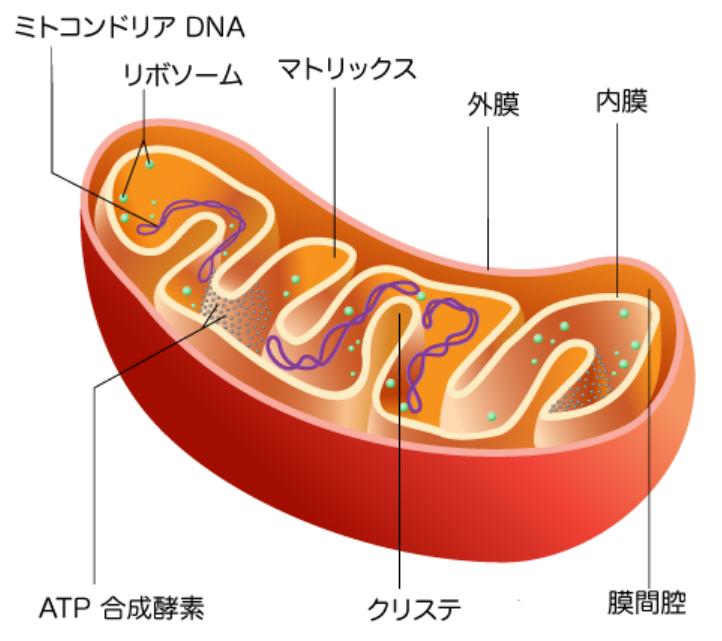
そこで、生物のエネルギー生成が行われている「細胞の熱力学的中枢」、『ミトコンドリア』の奥深く細胞呼吸の部位を観察することから始める。
ここでは、あなたが食べて残されたものの分子からプロトンが剥ぎ取られ、巨大な『呼吸鎖複合体』(注19)のうち最初で最大の構成要素である『複合体Ⅰ』に渡される。
(注19)「呼吸鎖複合体」とは、好気呼吸、嫌気呼吸関わらず細胞呼吸を行うほとんどの生物に見られる膜(ミトコンドリア内膜、チラコイド膜、原核生物の細胞膜)に存在する分子量10万から100万程度の巨大タンパク質である。「呼吸鎖複合体 I, II, III, IV」 からなり、ATP合成酵素は「呼吸鎖複合体 V」 とも呼ばれる。
この巨大な複合体Ⅰは45種類ものタンパク質から成る複雑な構造を有し、それぞれのタンパク質は数百のアミノ酸から構成されている。ATPが人間ほどの大きさとすれば、複合体Ⅰは摩天楼ほどにもなる。さらに、これらはただの巨大な建造物ではなく、蒸気機関のように働く巨大な動的なマシンなのである。
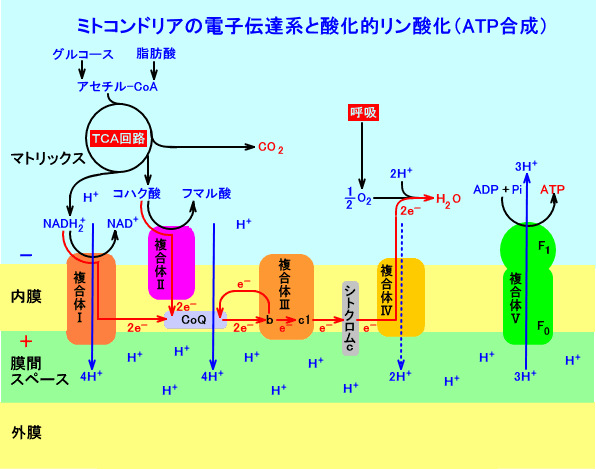
この複合体Ⅰには、電子がプロトンとは別に運ばれる。一方から取り込まれ、他方の膜の奥から排出される。その後、電子はさらに2つの巨大なタンパク質複合体を経由する。これらを合わせたものが『呼吸鎖』と呼ばれる。各複合体には複数の「レドックス中心」(複合体Ⅰにはおよそ9個)を含み、電子を一時的に保持する機能を有している。電子はこれらの「中心」から次の「中心」へと跳躍する。それらの「中心」が等間隔で散らばっている事実は、電子が『量子トンネル効果』を示していることを示唆している。電子の目から見えるのは、遠く離れていないかぎり、次のレドックス中心だけである。
ここでは、距離はオングストローム(Å)(1Åはほぼ水素原子1個のサイズ)を単位として測定される。どのレドックス中心も次のものとの間隔が14Å以内である。そして、前のレドックス中心よりわずかに電子への親和性が高い限り、電子はレドックス中心の連なる経路を跳躍して進む。これは、適度な間隔で並んだ飛び石をたどって川を渡るようなイメージに近い。
(注20)ニコチンアミドアデニンジヌクレオチド (英: nicotinamide adenine dinucleotide) とは、全ての真核生物と多くの古細菌、真正細菌で用いられる電子伝達体であり、エネルギー運搬体でもある。この物質は、さまざまな脱水素酵素の補酵素として機能し、酸化型 (略号NAD+) および還元型 (NADH、またはNADH2とも記す) の2つの状態を取り得る。二電子還元を受けるが、中間型は生じない。これらはATPを合成する解糖系、TCA回路、呼吸系などの一般的な代謝で利用される。
電子は3つの巨大な呼吸鎖複合体を一気に渡る。酸素が強く電子を引き寄せる力により、電子は前方へ引き寄せられる。これは遠隔作用とは異なり、電子が他の場所よりも酸素にくっついている確率の問題である。結局のところ、この現象はタンパク質と脂質によって隔てられ、食物から酸素へ電子を流す導線となる。この電流は、その場所で全てを動かすのである。電子は辿るべき経路に沿って跳躍し、酸素に到達するルートにだけ関心があって、周りのマシンは眼中にない。
しかし、これらの巨大なタンパク質複合体には多数の連動スイッチが存在している。電子がレドックス中心に存在するとき、隣接するタンパク質は特定の構造を取る。その構造は、電子が次のレドックス中心へ進むとわずかに変わり、負電荷の場所が移り、正電荷もそれに続き、弱い結合のネットワーク全体が調整しなおされて、巨大な組織が一瞬で新しい立体構造に変化する。タンパク質の一ヵ所で小さな変化が起こると、他のどこかで洞窟のようなチャネル(通路)が開く。すると新たに電子がやってきて、マシンが先ほどの状態に戻る。
このプロセスが1秒間に何十回も繰り返される。現在では、こうした呼吸鎖複合体の構造について、わずか数オングストロームというほぼ原子レベルの分解能で、多くのことが理解されている。タンパク質の電荷によってしかるべき場所に固定された水分子に、プロトンがどのようにして結びつくのかはわかっている。そうした水分子が、どのようにしてチャネル自体の構造変化の際に移動するのかもわかっている。
さらに、プロトンがどのように動的に開閉する隙間を素早く通り、水分子間でやり取りされるかも理解されている。プロトンが通過した途端に閉まることで、後戻りが防がれる。この巨大で精巧な運動機構は、ミトコンドリア内膜の片側からもう一方の片側へとプロトンを運ぶという一つの役割を果たす。
呼吸鎖の最初の複合体は、2個の電子が通過するごとに、4個のプロトンを膜を越えて送り出す。2個の電子はその後、第二の複合体(厳密には複合体Ⅲであり、複合体Ⅱは複合体Ⅰとは独立した経路を持つ)に進入し、この複合体はさらに4個のプロトンを膜を越えて送り出す。最後に、最終的な呼吸鎖複合体で、電子は酸素と出会うが、その前に2つのプロトンが更に膜を越えている。結果として、食物から抽出された2個の電子に対して、合計で10個のプロトンが膜を越えて輸送されている。
酸素へ向かう電子の流れが放出したエネルギーの半分弱が、プロトン勾配に貯えられる。そのパワーのすべて、その巧みさのすべて、その巨大なタンパク質の構造のすべてが、ミトコンドリアの内膜を越えてプロトンを汲み出すのにもっぱら向けられている。
ミトコンドリアの主な役割はプロトンの抽出であり、全体として毎秒1021個以上のプロトンを抽出している。この数は既知の宇宙に存在する恒星の数に近い。
1つのミトコンドリアには、数万個の呼吸鎖複合体が存在する。1つの細胞には、数百から数千個のミトコンドリアが含まれている。あなたの体の40兆個の細胞には、最少でも1000兆個のミトコンドリアが存在し、その複雑に入り組んだ膜の表面積は合計で約14000平方メートルになる。これはサッカー場4個分の広さに相当する。
しかし、これはミトコンドリアの仕事の半分に過ぎない。残りの半分は、そのエネルギーを利用してATPを生成することである。
ミトコンドリアの膜はプロトンをほぼ通さず、プロトンが通過した直後には動的なチャネルがすぐに閉じる。プロトンは非常に小さく(水素の原子核に過ぎない)、そのサイズのために、プロトンを遮断するのは容易なことではない。プロトンは水分子を即座に通過するため、膜は水が侵入しないように完全に密封されている必要がある。さらに、プロトンは正の電荷を持つ。
密閉された膜を越えてプロトンを汲み出すと、二つのことがなし遂げられる。
- 第一に、膜の両側でプロトン濃度(水素イオン濃度)に差が生じる。
- 第二に、電荷の差も生み出し、外側が内側に比べ正になる。すると、膜をはさんで150~200ミリボルト程度の電気化学ポテンシャルが生じる。膜は非常に薄い(厚さ6ナノメートルほど)ため、この電荷の差は短い距離でとてつもなく大きいものとなる。膜のそばで感じる電場の強度、電界の強さは3000万V/mで、これは稲妻に匹敵する。
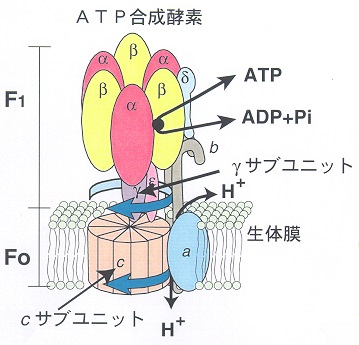
この大きな電気化学ポテンシャルは、『プロトン駆動力』と呼ばれ、最も壮大なタンパク質ナノマシンと言える『ATP合成酵素』(図34~36)を駆動する。駆動とは運動のことであり、ATP合成酵素は実のところ回転モーター(F0モーター)で、プロトンの流れがクランクシャフトを回し、それが触媒部位であるヘッド(触媒ヘッド、F1モーター)を回す。この機械的な力がATPの合成を進行させる。たとえて言えば、このタンパク質は水力発電のタービンの役割を果たし、それによって、膜の障壁の向こうにある貯水池に閉じ込められていたプロトンが、流れ下る水のようにタービンに押し寄せ、回転モーターを回す。
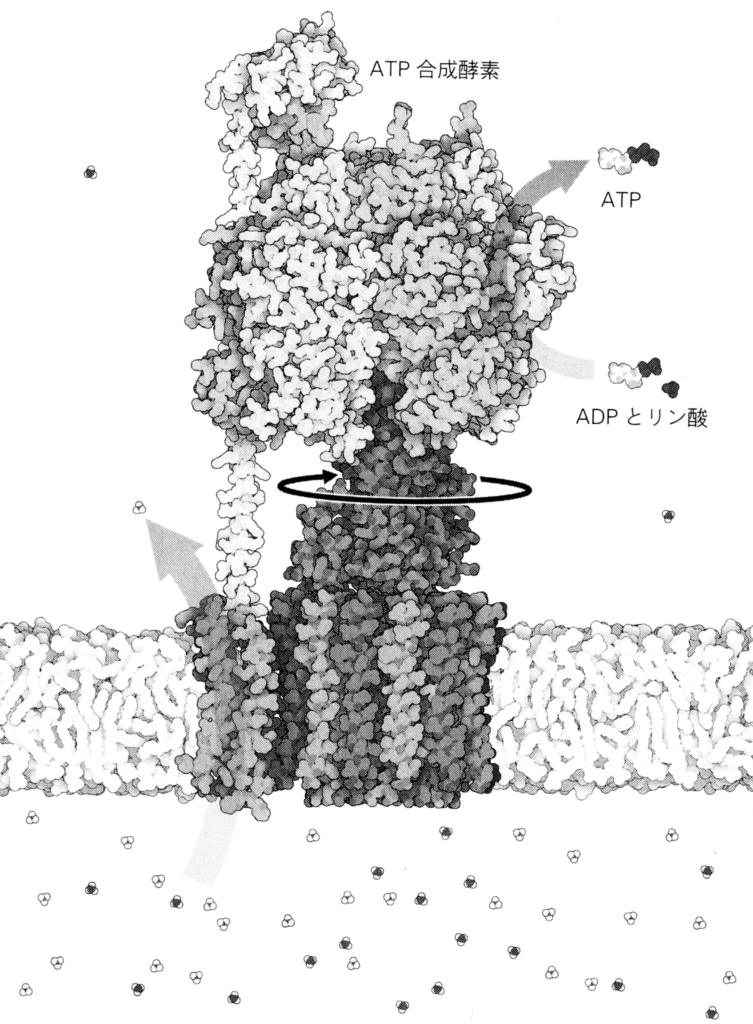
その作用機構はまだ完全には理解されていない。
- 個々のプロトンがどのように膜内のCリング(Cサブユニット、モーター部分を構成する回転子)に結びつくのか、
- この回転するリングがどのようにクランクシャフトをねじり、触媒ヘッドの構造変化をもたらすのか、
- このヘッドで開閉する空隙がどのようにADPとPiをつかんで機械的に結合させ、新たなATPを生成するのか、
などについての理解は未完全である。これは最高レベルの精密ナノ工学と言える魔法の装置(図36)で、知れば知るほど驚かされる。
ATP合成酵素では、プロトンが10個通過する度に、回転するヘッドが全周し、新たに合成された3つのATP分子が基質(酵素によって化学反応を触媒する物質)として放出される。ヘッドは、毎秒100回以上の速さで回転する可能性がある。
ATPは生命の普遍的な『エネルギー通貨』と呼ばれているが、同様に『ATP合成酵素』や『プロトン駆動力』も、生命全体の進化史を通じて普遍的に保持されているのである。これらは遺伝コードと同様に普遍的な存在である。それゆえに、
- 『ATP合成酵素』は『DNAの二重らせん』と同様に、生命にとって象徴的な存在である
と位置づけられる。
『プロトン駆動力』の概念は、20世紀に生体エネルギー論の分野で革命を起こした科学者の一人、ピーター・ミッチェルによるものである。彼は、1961年に『化学浸透圧説』を発表したが、『化学浸透圧(chemiosmotic)』は、膜を越えてのプロトンの移動を指すミッチェルの表現である。この場合の「osmotic(浸透圧)」という言葉は、「押す」という意味で、一般的に理解されている「半透膜を水が通り抜ける」という用法とは異なる。
すなわち、『呼吸』はプロトンを濃度勾配に逆らって薄い膜を越えて押し出すために、ミッチェルはchemiosmotic(化学+押す)という表現を用いたのである。つまり、呼吸は、20世紀半ばまで考えられていたような反応媒介物による化学反応ではなく、
- 電子の流れをATP合成に結びつけるメカニズムは、実際には半透膜を隔てて生じる『プロトン勾配-プロトン駆動力-』なのである。
基本的にすべての生命は、『レドックス反応』を利用して、膜をはさんだプロトン勾配を生み出している。いったいなぜ生命は、そのようなメカニズムに依存するのだろうか。すなわち、
- なぜ生体細胞はすべて、「レドックス反応」を自由エネルギーの源として利用するのか、
さらに深く考えると、
- なぜ「電子」なのか、なぜ「プロトン」なのか
という疑問に至る。
そこで、この二つの基本的な疑問について考察を進めることにしたい。
生命は結局のところ電子
では、いったいなぜ生命はレドックス反応を利用するのだろうか。
私たちが知っている生命の基盤は『炭素』で、具体的には「部分的に還元された形態の炭素」である。窒素やリンなどの元素も比較的少量必要ではあるが、極端な近似で、「生命の化学式」は『CH2O』であると言える。二酸化炭素を起点に考えると、生命は、水素(H2)などからCO2への電子やプロトンの移動を必要とする。理論的には、電子の供給源は何でも良い。水(H2O)、硫化水素(H2S)、さらには第一鉄(Fe2+)から電子を奪うことも可能である。つまり、
- 電子がCO2へ運ばれることが重要であり、それらの移動はすべてレドックス反応なのだ。ちなみに、「部分的に還元された」という表現は、CO2がメタン(CH4)まで完全には還元されないことを指す。
もし生命が炭素以外の元素を利用できたとしたらどうだろうか。他の元素、たとえば『ケイ素(シリコン)』なども考えられるだろう。しかし、炭素には多くの利点がある。すなわち、
- 炭素原子は一つで四つの強力な結合を形成でき、その結合は化学的に近いケイ素が形成するものより遥かに強い。
- この結合によって、多種多様な長鎖の分子、特にタンパク質、脂質、糖、DNAを作り出すことが可能である。
- ケイ素では、このような多様な化学反応を達成することは難しい。また、ケイ素で二酸化炭素に匹敵する気体の酸化物は存在しない。
CO2をレゴのブロックのようなものと考えてみると、
- それは空気から取り出され、一度に一つの炭素を他の分子に加えることができる。
- 一方で酸化ケイ素は、砂で建築をしようとするようなものである。ケイ素などの元素はわれわれ人間のような比較的高等な知的生命が利用するには適しているかもしれないが、ケイ素を使って一から生命を形成する可能性は、なかなか見えてこない。
また、確率と予測性の問題として、可能性は圧倒的に低いように見える。はるかに優れているほかに、炭素は宇宙全体にはるかに豊富に存在もする。従い、ざっくり言って、生命は炭素ベースであるべきなのである。
そこで、今度は問題を逆から考えてみよう。具体的に、「レドックス反応」がなぜ有用であるのかを探求するのである。
「呼吸」について考えるとき、人間だけでなく、他の生命体も考慮に入れるべきである。私たちは食物から電子を奪取し、呼吸鎖を通じて酸素に運んでいるが、
ここでの重要なポイントは、電子の供給源と呼吸場所の両方を異なるものに置き換えることも可能であるということである。
食物を酸素の中で燃焼させると、エネルギーの収集効率は最大となるが、その根底にある原理は、非常に多様で柔軟である。事実、必ずしも有機物を摂取する必要はないのである。「水素ガス」、「硫化水素」、「第一鉄」は、前述したようにすべて電子供与体であり、反対側に十分強力な酸化剤が存在すれば、自身の電子を呼吸鎖に渡すことが可能である。それ故に細菌は、私たちが呼吸に使用するのと基本的に同じタンパク質装置を使って、岩石や鉱物やガスを「食べる」ことができるのである。
酸素の存在も必須ではない。他の多くの酸化剤が、ほぼ同様の作業を遂行できる。「硝酸塩」や「亜硝酸塩」、「硫酸塩」、「亜硫酸塩」など、そのリストは無限に続く。これらの酸化剤(酸素と類似した反応性を示すためにこのように称される)は全て、食物などの供給源から電子を吸収する能力を持つ。細菌は岩石を「食べる」だけでなく、「呼吸する」ことも可能である(注21)。
(注21)生物が呼吸によってエネルギーを獲得する場合、有機物などの代謝に伴って放出された電子が酸素などの電子受容体に受け渡される。この際、電子供与体(代謝によって生み出された『NADH』(注22)など)と電子受容体の酸化還元電位の差に相当するエネルギーが放出され、生物は主にこのエネルギーを利用してATPを合成している。酸素以外の物質を電子受容体として用いる呼吸を『嫌気呼吸』といい、酸素のない環境に生息する細菌または古細菌(『嫌気性菌』)は硝酸、硫酸、二酸化炭素などの物質を電子受容体として利用している。嫌気性細菌の中には酸化鉄などの固体の金属化合物を電子受容体として呼吸を行うものもおり(『鉄還元菌』)、このような細菌は細胞外に存在する固体に電子を伝達するための経路(細胞外電子伝達経路)を備えている。
(注22)『NADH(ニコチンアミドアデニンジヌクレオチド)』とは、全ての真核生物と多くの古細菌、真正細菌で用いられる電子伝達体である。さまざまな脱水素酵素の補酵素として機能し、酸化型および還元型の2つの状態を取り得る。二電子還元を受けるが、中間型は生じない。略号であるNAD⁺のほうが論文や口頭でも良く使用されている。
電子供与体と受容体の利用に関するこの多様性は、それらの多くの反応が遅いことに助けられている。あらゆる生化学的メカニズムは自発的に生じるもので、必ず反応性の高い環境に駆り立てられる必要がある。だが、環境の反応性があまりにも高いと、すぐに反応が進み、生命活動の原動力となる自由エネルギーが何も残らなくなる。
たとえば、大気はフッ素ガスでいっぱいにはならないだろう。すぐに何とでも反応して消滅してしまうからである。しかし多くの物質は、反応がとても遅いため、自然の熱力学的平衡をはるかに超えるレベルまでたまる。機会さえ与えられれば、酸素は有機物と激しく反応してこの惑星の何もかもを燃やしてしまうだろうが、この乱暴な傾向は、地球を莫大な年月にわたり安定させている幸運な化学的状況によって和らげられている。メタンや水素などのガスは、有機物よりもさらに激しく酸素と反応するが、それらの反応にも速度論的障壁があるので、こうしたガスはどれも『動的不均衡』の状態で何年も空気中に共存できる(注23)。同じことは、硫化水素から硝酸塩まで、ほかの多くの物質にも言える。それらは強制的に反応させることができ、そのときには大量のエネルギーが放出され、生体細胞に利用できるが、しかるべき触媒がないとたいしたことは起きない。
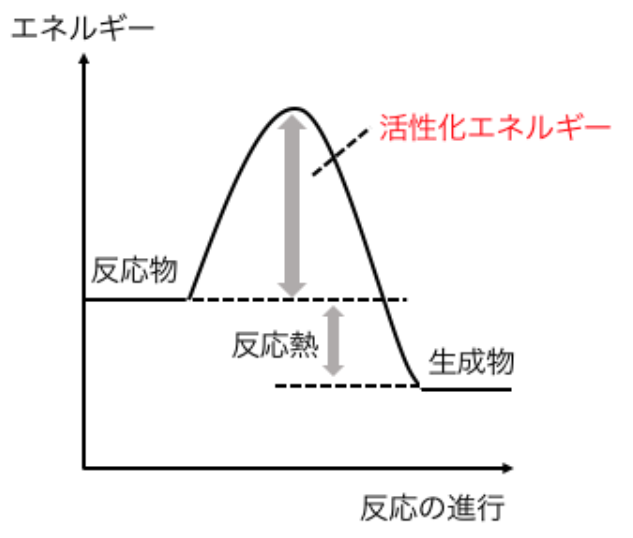
(注23)2H2 + O2 → 2H2O という反応は発熱反応であり、自由エネルギーも減少するから、自発的に起こる反応である。しかし、H2とO2を混ぜただけでは反応は起こらず、温度をかなり高くしないと起こらない。これは次のように考えられている。反応を起こすには「エネルギー障壁」(図27)があり、通常、分子はこのエネルギー障壁を乗り越えることができない。しかし、温度が上がると分子の中には、障壁を乗り越えるのに十分なエネルギーを持つものが現れてきて、反応が進行するようになる。このエネルギー障壁を『活性化エネルギー』とよぶ。 nA+ mB→Cの場合、一般に反応速度v は以下のように書ける場合が多い。
v= k[A]n[B]m
ここで、kを『反応速度定数』という。なお、反応速度定数kと活性化エネルギーEaおよび温度Tには、次の重要な関係
k=αe-Ea/RT
がある。
生命はこのような『速度論的障壁』を利用しながら、エントロピーをより速く増大させている。こうした点で生命を『エントロピー発生装置』と定義する人さえいる。ともあれ、
- 生命はまさに速度論的障壁があるがために存在している、その障壁を乗り越えることを得意としているのである。
- 速度論的障壁の向こうに抑え込まれている高い反応性という抜け道がなければ、生命がそもそも存在できたかどうかも疑わしい。
しかも、
- 多くの電子供与体と受容体が水溶性にて化学的に安定で、たやすく細胞に出入りするという事実は、熱力学が求める反応性の高い環境を確実に細胞内へ、細胞の重要な膜へ持ち込めることを意味する。
- そのおかげで「レドックス反応」は、熱エネルギーや力学的エネルギー、紫外線放射や稲妻よりも、生物学的に有用なエネルギーの流れの一形態としてはるかに扱いやすくなっている。
意外にも、呼吸は光合成の基礎でもある。光合成には複数のタイプが存在し、各々において、日光のエネルギーを光子として吸収した色素(通常は葉緑素)が電子を励起し、レドックス中心の連鎖へ送り、受容体(この場合は二酸化炭素)まで達成させる。
電子を失った色素は、近くにある供与体-水であれ、硫化水素であれ、第一鉄であれ-から電子を補給する。呼吸の場合と同様、電子供与体の種類は原理上問題ではない。「酸素非発生型」の光合成は硫化水素や鉄を電子供与体として使い、硫黄や錆びた鉄の沈殿物を廃物として残す。一方、「酸素発生型」の光合成は、より困難な供与体-水-を用い、酸素を廃物として排出する。
だが重要なのは、こうした種々のタイプの光合成がすべて、
- 明らかに呼吸に由来しているという事実である。
- 図37に示すように、まったく同じ呼吸系タンパク質、同じタイプのレドックス中心、膜を挟んでの同じプロトン勾配、同じATP合成酵素、を用いている。
- 唯一の違いは色素の変化で、『葉緑素』は、多くの太古の呼吸系タンパク質で使われていた『ヘム』(注24)という色素と密接に関係している。太陽エネルギーの利用は世界を変えたが、分子の観点では呼吸鎖に電子が速く流れるようにしただけだった。
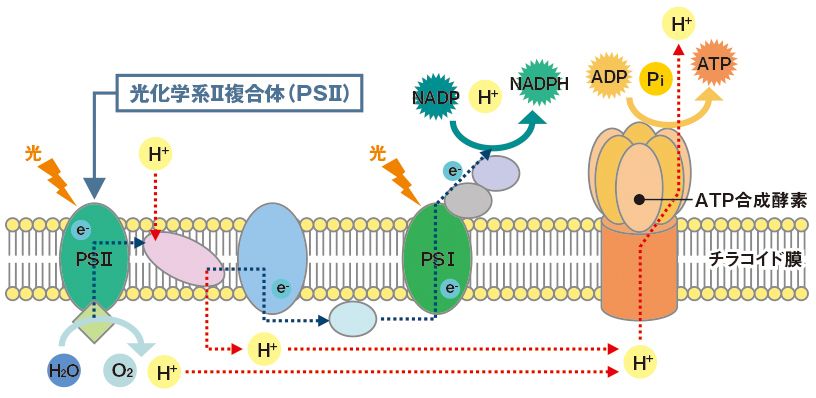
(注24)「葉緑体」は、日光を受けると、根からとり入れた水と空気中の二酸化炭素を使ってでんぷんを作るとともに、酸素を放出する。この時、「葉緑素」は日光のエネルギーを吸収する役割を担う。このようなはたらきを「光合成」という。
その際「ヘム」は葉緑体でアポ蛋白質と結合することにより、電子伝達反応、酸化還元反応、活性酸素消去など様々な機能を果たしている。なお、「ヘム」は中心金属として鉄を持つ『ポルフィリン錯体』(ポルフィンおよびその誘導体ポルフィリンを配位子とする、有色の金属錯体の総称。ポルフィリンは大環状平面分子で、中心にある4個のN原子で金属原子と結合する)で、酸化還元によって鉄が二価と三価を行き来する性質や二価鉄に酸素分子が結合する性質を利用することによって、電子伝達、酸素の運搬、酵素の活性中心として用いられる。
したがって、
- 呼吸の大きな利点は途方もない多様性である。ほぼどんなレドックス対も(どんな電子供与体と電子受容体のペアも)、電子を呼吸鎖に流すのに利用できる。
アンモニウムイオンから電子を取り出すタンパク質は、硫化水素から電子を取り出すタンパク質とわずかに違うが、主眼は同じで密接なつながりを持つバリエーションなのである。また、呼吸鎖の反対側の末端でも、電子を硝酸塩や亜硝酸塩に渡すタンパク質は、電子を酸素に渡すタンパク質と違うが、どれも関連している。互いに十分に似ているので、取り替えて利用することもできる。
これらのタンパク質は共通のオペレーティング・システムに接続できるため、任意の環境に適応させることが可能である。ただ原理的に互換性があるだけでなく、実際には様々な箇所で自由に交換される。ここ数十年の間に、遺伝子の水平移動(細胞間で小銭のように遺伝子の小さなカセットを渡し合うこと)が細菌や古細菌で頻発していることがわかってきた。呼吸系タンパク質をコードしている遺伝子群は、その中でもとりわけよく水平移動で交換される。それらをまとめて、生化学者のヴォルフガング・ニッチケは『レドックスタンパク質構築キット』と呼んでいる。
- これらの要素をすべて考慮すると、レドックス反応は宇宙のどの生命にとっても重要であると言えるだろう。
- 他の形態のエネルギー源も考えられるが、炭素を還元するレドックス反応の必要性と、呼吸の多くの利点を考えると、地球上の生命がレドックス反応をエネルギー源として用いるのは驚くべきことではない。
しかし、呼吸の本質的なメカニズムである、膜を挟んでの『プロトン勾配』は、まったく別の問題である。
- 呼吸系タンパク質が遺伝子の水平移動によって様々な場所に分散され、どの環境でも機能するように混ぜ合わせられるのは、共通のオペレーティング・システムが存在するという事実に由来している。
- しかし、生命がなぜプロトンを利用するのか、プロトン勾配がなぜレドックス反応に関与すべきなのかという明確な理由が解明されていない限り、地球上や宇宙の他のどこかに存在する可能性のある「生命の特質」について予測することは困難であるだろう。
生命は結局のところプロトン
『化学浸透共役』(注25)の進化は未だに謎のベールに包まれている。あらゆる生命が化学浸透圧を利用しているという事実は、化学浸透共役が進化の極めて初期段階で発生した可能性を示唆している。もし後から出現したとすれば、なぜどのようにして普遍的になったのか、なぜプロトン勾配がほかのすべてにすっかり取って代わったのか、を説明しにくいはずだからである。ここまで普遍的に見られる仕組みは極めて稀である。
(注25)細胞内で、「ATPの化学合成」と、「電子伝達系のエネルギーを利用したプロトンの膜透過(浸透)」が組になってエネルギー変換を行う機構を指す。
ここで、『化学浸透』とは、イオンが電気化学的勾配に従って半透膜を越えて移動することを意味する。細胞呼吸や光合成の過程において、膜を越えた水素イオン(H+)の移動によって「アデノシン三リン酸(ATP)」が産生されることなどが挙げられる。水素イオン(プロトン)は高濃度の領域から低濃度の領域へと拡散し、膜を挟んだプロトンの電気化学的勾配がATPの合成に利用される。この過程は膜を越えて水が拡散する「浸透」と関連しており、そのため『化学浸透』と呼ばれている。ATP合成酵素は、化学浸透を利用してATPを合成する酵素である。ATP合成酵素は膜を越えてプロトンの通過を可能にし、その自由エネルギー差を利用してアデノシン二リン酸(ADP)をリン酸化してATPを合成する。化学浸透によるATP産生は、ミトコンドリアと葉緑体とともに大部分の細菌と古細菌でも起こる。
一つの例として、「細菌」と「古細菌」をとり挙げてみよう。すべての生命は『遺伝コード』を共有しており、いくつかの基本的な情報処理も普遍的に保存されている。たとえばあらゆる生体細胞で、『DNA』は『RNA』に転写され、それが『リボソーム』というナノマシンでタンパク質に物理的に翻訳される。ところが『原核生物』(細胞内にDNAを包む核を持たない単細胞生物)の二大ドメインである『細菌』と『古細菌』(注26)との差異は非常に大きい。
(注26)生物の主要な系統の一つである。『細菌(バクテリア)』、『真核生物(ユーカリオタ)』と共に、全生物界を3分している。『古細菌』は形態や名称こそ細菌と類似するが、細菌とは異なる系統に属しており、その生態機構や遺伝子も全く異なる。非常に多様な生物を含むが、その代表例として「高度好塩菌」、「メタン菌」、「好熱菌」などが良く知られている。
物理的な外観では、細菌と古細菌を区別するのは困難だが、多くの生化学的・遺伝学的特徴において、これら二つのドメインは全く異なっている。例えば、『遺伝コード』と同じぐらい生命にとって基本的なものと考えられる『DNA複製』の具体的なメカニズムは、必要なほぼすべての酵素も含め、細菌と古細菌ではまったく異なることがわかっている。また、『細胞壁』(植物や菌類、細菌類の細胞に見られる構造で、動物細胞には存在しない。細胞膜の外側に位置するために細胞外マトリクスの一部をなす)も、細菌と古細菌では化学的にまるで異なるし、発酵の生化学的経路も異なっている。化学浸透共役に必須である「細胞膜」さえ、細菌と古細菌では生化学的に異なっている。つまり、細胞の内側と外側を隔てる障壁や遺伝物質の複製は、根本的に保存されてはいないのである。これら以上に細胞の生命にとって重要なものはないはずにもかかわらず。
上記はきわめて根本的な差異で、細菌・古細菌双方のグループの共通祖先について、深刻な疑問をもたらす。共通の形質は共通の祖先から受け継がれるが、異なる形質はふたつの系統で独立に生じたのだとすれば、その祖先はどのような細胞だったと考えられるのだろうか。
DNA転写、リボソームによる翻訳、ATP合成酵素、そして部分的なアミノ酸の生合成は存在していたにせよ、それ以上の共通点は二つのグループ間でほとんど見られないのである。このような差異が多い中で、『化学浸透共役』のみが普遍的なのである。
しかし、一方では、化学浸透共役が進化の非常に早い段階で生じていたという考えには、いくつかの困難な問題が存在している。
ひとつは、メカニズムの純然たる複雑さである。すでに私たちは、巨大な呼吸鎖複合体とATP合成酵素(ピストンと回転モーターをもつ途方もない分子マシン)でその経験は積んではきたが、これらはDNA複製が登場する以前、進化の最初期に、生み出され得たのかとかということに疑問が呈されている。
第二の問題は、膜そのものであり、早い時期におそるべき複雑化をなし遂げたという問題がある。現代の細胞では、化学浸透共役は、膜がほとんどプロトンを通さない場合にのみ働く。しかし初期の膜と考えられるものは、それを用いたあらゆる実験から、プロトンをよく通していたことがうかがえる。実は、プロトンを通さないようにするというのはきわめて難しいのである。問題というのは、いくつもの複雑化したタンパク質がプロトンを通さない膜に埋め込まれるまで、化学浸透共役が役に立たないように見えるという点にある。膜にタンパク質が埋め込まれて初めて、それは役に立つのである。ならば、いったいどうしてこれらのパーツが先に進化を遂げたのだろうか。
地球上の「生命の進化」については、従来から大きな疑問がいくつも指摘されてきた。詳細は別の機会に譲るとし、その疑問群を挙げると、
- 生命はなぜこんなにも早く誕生したのか
- なぜ数十億年にわたり形態学的複雑さが停滞したのか
- なぜ複雑な真核細胞は40億年間でただ一度しか生じていないのか
- なぜすべての真核生物は、有性生殖やふたつの性別から老化まで、細菌や古細菌には見られない不可解な形質を多く持っているのか
というものである。ここに、これらの疑問に同じくらい気がかりな疑問がもう二つ、
- あらゆる生命は、なぜ、膜を介してのプロトン勾配という形でエネルギーを保存しているのか
- この奇妙だが基本的なプロセスはどのように、また、いつ進化を遂げたのか
が加わる。実際、これら二つの疑問は、全体として互いに関連し合っているのであるが。
そこで、次章にて、これらの疑問に答えながら、
- 自然に生じたプロトン勾配が、非常に特異な環境ではあるが、宇宙ではほぼ間違いなく普遍的な環境で、生命を誕生させた。その環境に必要なのは、岩石と、水と、CO2だけであった。
さらに、
- 化学浸透共役が地球上の生命の進化を、数十億年にわたり細菌と古細菌の複雑さまでに制約した。
- 1個の細菌がなぜか別の1個の中に入るというただ一度の出来事が、こうした細菌に絶えず加わっていた強い制約を乗り越えさせた。その内部共生で生まれた真核生物は、桁違いに大きなゲノムをもち、それが形態上の複雑さの原材料となった。
- 宿主細胞とその内部共生体(ミトコンドリアとなるもの)との密接な関係が、真核生物に共通する多くの不可解な特性の背後にあった。
などということを解き明かしていこう。
進化は、宇宙のどこでも、同じような制約に導かれて同じような道筋に沿って展開される傾向があると考えられる。
図表
図25 枯草菌の芽胞
名古屋学芸大学、管理栄養学部、なるほど豆知識
(https://nutrition.nuas.ac.jp/tips/000037.html)
図26 脂質二重層とリン脂質分子の例
『ウィキワンド(Wikiwand)』「リン脂質」
(https://www.wikiwand.com/ja/%E3%83%AA%E3%83%B3%E8%84%82%E8%B3%AA)
図27 エネルギー障壁(活性化エネルギー)
受験メモ・化学・反応速度とは?活性化エネルギーも詳しく解説!
(http://www.jukenmemo.com/chemistry/theory/reaction-rate/)
図28 ATPの分解と合成
受験生学習支援ブログ
(https://wearewhatwerepeatedlydo.com/bio-atp/)
図29 酸素が鉄を酸化する反応
金沢大学・理工学域自然システム学類佐藤研究室(環境鉱物学研究室)ホームページ
(http://earth.s.kanazawa-u.ac.jp/Environmental_Mineralogy/sato/geohist_8/sld002.html)
図30 呼吸によるATPの合成とエネルギーの放出
NHK高校講座、生物基礎、第6回 呼吸
(https://www.nhk.or.jp/kokokoza/tv/seibutsukiso/archive/resume006.html)
図31 代表的な鉄硫黄(Fe-S)クラスター
埼玉大学理学部分子生物学科・分子統御研究室、研究内容「鉄硫黄クラスターと鉄硫黄タンパク
質」
(http://park.saitama-u.ac.jp/~tougyo/yan_jiu_nei_rong.html)
図32 ミトコンドリアの構造
生命系のための理工学基礎・生化学・ミトコンドリアとは
(https://rikei-jouhou.com/mitochondria/)
図33 ミトコンドリアの電子伝達系と酸化的リン酸化(ATP合成)
脂質と血栓の医学・生化学の知識・電子伝達系と酸化的リン酸化
(http://hobab.fc2web.com/sub2-respiratory-chain.htm)
図34 ミトコンドリアとATP合成酵素
吉田賢右「生き物のエネルギー通貨を生み出すナノモーター—ATP合成酵素の回転運動を世界で
初めて観察—」京都産業大学 研究・社会連携 サイエンス&テクノロジー
(https://www.kyoto-su.ac.jp/project/st/st11_06.html)
図35 ATP合成酵素の構成図
科学技術振興事業団、お知らせ・平成11年11月25日
(https://www.jst.go.jp/pr/announce/19991126/index2.html)
図36 ATP合成酵素
「生命、エネルギー、進化THE VITAL QUESTION Why Is Life the Way It Is?」
ニック・レーン Nick Lane著、斉藤隆央訳、株式会社 みすず書房、2016年9月13日 第1刷、
2018年3月9日 第10刷 図10「ATP合成酵素の構造」、85頁
図37 光合成の明反応の流れ
国立研究開発法人科学技術振興機構、事業成果、環境エネルギー、光合成最大の謎を解明
(https://www.jst.go.jp/seika/bt3-4.html)
